Home
- Introduction
- Installation of the HHsuite and its databases
-
Brief tutorial to HHsuite tools
- Overview of programs
- Searching databases of HMMs using HHsearch and HHblits
- Generating a multiple sequence alignment using HHblits
- Example: Comparative protein structure modeling using HHblits and MODELLER
- Visually checking an MSA for corrupted regions
- Building customized databases
- Example: Building a database from the PDB
- Modifying or extending existing databases
-
Frequently asked questions
- How do I report a bug?
- What are HMM-HMM comparisons and why are they so powerful?
- When can the HH-suite be useful for me?
- What does homology mean and why is it important?
- How can I verify if a database match is homologous?
- What does the maximum accuracy alignment algorithm do?
- How is the MSA diversity Neff calculated?
- Do HHsearch and HHblits work fine with multi-domain sequences?
- How do I best build HMMs for integral membrane proteins?
- How do I reconcile overlapping and conflicting domain predictions?
- How can I build a phylogenetic tree for HMMs?
- Can I weight some residues more than others during the HMM-HMM alignment?
- Don’t I need to calibrate my query or database HMM?
- Should I use the -global option to build MSAs if I am intersted in global alignments to these database HMMs?
- How many iterations of hhblits should I use to search my customized genome database?
- How can I retrieve database A3M or HHM files from my HH-suite database?
- How can I build my own UniProt database for HHblits?
- Will you offer an HHsuite database that includes environmental sequences?
- How do I read the sequence/HMM headers in the pdb70 databases?
- Why do I get different results when reversing the roles of query and template?
- Why do I sometimes get the same database hit twice with different probabilities?
- Why do I get different results using HHblits and HHsearch when I search with the same query through the same database?
- Why do I get different results when I perform 2 and 1+1 iterations with HHblits?
- Why do I get a segmentation fault when calling hhblits on our new machine?
- I obtain different results for the same query sequence on the web server and on my local version using HHblits and HHsearch.
- I find an alignment of my query and a template with the exact same sequence, but they are aligned incorrectly.
- HHsearch/HHblits output: hit list and pairwise alignments
- File formats
-
Summary of command-line parameters
- hhblits – HMM-HMM-based lighting-fast iterative sequence search
- hhsearch – search a database of HMMs with a query MSA or HMM
- hhfilter – filter an MSA
- hhmake – build an HMM from an input alignment
- hhalign – align a query MSA/HMM to a template MSA/HMM
- reformat.pl – reformat one or many alignments
- addss.pl – add predicted secondary structure to an MSA or HMM
- hhmakemodel.pl and hhmakemodel.py – generate MSAs or coarse 3D models from HHsearch results file
- hhsuitedb.py – Build an HHsuite database
- splitfasta.pl – Split multi-sequence FASTA file into single-sequence files
- renumberpdb.pl – Renumber indices in PDB file to match input sequence indices
- cif2fasta.py – Create a fasta file from cif files
- pdbfilter.py – Filter sequences from the PDB (requires MMseqs2)
- License
- Contributors and Contact
- Acknowledgments
The HH-suite is an open-source software package for sensitive protein sequence searching based on the pairwise alignment of hidden Markov models (HMMs). It contains HHsearch and HHblits among other programs and utilities. HHsearch takes as input a multiple sequence alignment (MSA) or profile HMM and searches a database of HMMs (e.g. PDB, Pfam, or InterPro) for homologous proteins. HHsearch is often used for protein structure prediction to detect homologous templates and to build highly accurate query-template pairwise alignments for homology modeling. In the CASP9 competition (2010), a fully automated version of HHpred based on HHsearch and HHblits was ranked best out of 81 servers in template-based structure prediction. HHblits can build high-quality MSAs starting from single sequences or from MSAs. It transforms these into a query HMM and iteratively searches through Uniclust30 database by adding significantly similar sequences from the previous search to the updated query HMM for the next search iteration. Compared to PSI-BLAST, HHblits is faster, up to twice as sensitive and produces more accurate alignments. HHblits uses the same HMM-HMM alignment algorithms as HHsearch, but it employs a fast prefilter that reduces the number of database HMMs for which to perform the slow HMM-HMM comparison from tens of millions to a few thousands.
References:
-
Steinegger M, Meier M, Mirdita M, Vöhringer H, Haunsberger S J, and Söding J (2019) HH-suite3 for fast remote homology detection and deep protein annotation, BMC Bioinformatics, 473. doi: 10.1186/s12859-019-3019-7
-
Steinegger M, Meier M, Mirdita M, Vöhringer H, Haunsberger S J, and Söding J (2019) HH-suite3 for fast remote homology detection and deep protein annotation, bioRxiv, 560029. doi: 10.1101/560029
-
Mirdita M, von den Driesch L, Galiez C, Martin M J, Söding J, and Steinegger M (2016) Uniclust databases of clustered and deeply annotated protein sequences and alignments, Nucleic Acids Res., 45, D170–D176. doi: 10.1093/nar/gkw1081
-
Remmert M, Biegert A, Hauser A, and Söding J (2011) HHblits: Lightning-fast iterative protein sequence searching by HMM-HMM alignment. Nat. Methods 9, 173-175. doi: 10.1038/nmeth.1818
-
Angermüller C, Biegert A, Söding J (2014) Discriminative modelling of context-specific amino acid substitution probabilities. Bioinformatics 28 (24), 3240-3247. doi: 10.1093/bioinformatics/bts622
-
Söding J (2005) Protein homology detection by HMM-HMM comparison. Bioinformatics 21, 951-960. doi: 10.1093/bioinformatics/bti125
The HH-suite is an open-source software package for highly sensitive sequence searching and sequence alignment. Its two most important programs are HHsearch and HHblits. Both are based on the pairwise comparison of profile hidden Markov models (HMMs).
Profile HMMs are a concise representation of multiple sequence alignments (MSAs). Like sequence profiles, they contain for each position in the master sequence the probabilities to observe each of the 20 amino acids in homologous proteins. The amino acid distributions for each column are extrapolated from the homologous sequences in the MSA by adding pseudocounts to the amino acid counts observed in the MSA. Unlike sequence profiles, profile HMMs also contain position-specific gap penalties. More precisely, they contain for each position in the master sequence the probability to observe an insertion or a deletion after that position (the log of which corresponds to gap-open penalties) and the probabilities to extend the insertion or deletion (the log of which corresponds to gap-extend penalties). A profile HMM is thus much better suited than a single sequence to find homologous sequences and calculate accurate alignments. By representing both the query sequence and the database sequences by profile HMMs, HHsearch and HHblits are more sensitive for detecting and aligning remotely homologous proteins than methods based on pairwise sequence comparison or profile-sequence comparison.
HHblits can build high-quality multiple sequence alignments (MSAs) starting from a single sequence or from an MSA. Compared to PSI-BLAST, HHblits is faster, finds up to two times more homologous proteins and produces more accurate alignments. It uses an iterative search strategy, adding sequences from significantly similar database HMMs from a previous search iteration to the query HMM for the next search. Because HHblits is based on the pairwise alignment of profile HMMs, it needs its own type of databases that contain multiple sequence alignments and the corresponding profile HMMs instead of single sequences. The HHsuite database Uniclust30 is generated regularly by clustering the UniProt KB into groups of similar sequences alignable over at least 80% of their length and down to ∼30% pairwise sequence identity. This database can be downloaded together with HHblits. HHblits uses the HMM-HMM alignment algorithms in HHsearch, but it employs a fast prefilter that reduces the number of database HMMs for which to perform the slow HMM-HMM comparison from tens of millions to a few thousands. At the same time, the prefilter is sensitive enough to reduce the sensitivity of HHblits only marginally in comparison to HHsearch.
By generating highly accurate and diverse MSAs, HHblits can improve almost all downstream sequence analysis methods, such as the prediction of secondary and tertiary structure, of membrane helices, functionally conserved residues, binding pockets, protein interaction interfaces, or short linear motifs. The accuracy of all these methods depends critically on the accuracy and the diversity of the underlying MSAs, as too few or too similar sequences do not add significant information for the predictions. As an example, running the popular PSIPRED secondary structure prediction program on MSAs generated by HHblits instead of PSI-BLAST improved the accuracy of PSIPRED significantly even without retraining PSIPRED on the HHblits alignments.
HHsearch takes as input an MSA (e.g. built by HHblits) or a profile HMM and searches a database of HMMs for homologous proteins. The PDB70 database, for instance, consists of profile HMMs for a set of representative sequences from the PDB database; the scop70 database has profile HMMs for representative domain sequences from the SCOP database of structural domains; the Pfam domain database is a large collection of curated MSAs and profile HMMs for conserved, functionally annotated domains. HHsearch is often used to predict the domain architectures and the functions of domains in proteins by finding similarities to domains in the pdb70, Pfam or other databases.
In addition to the command line package described here, the interactive web server runs HHsearch and HHblits. It offers extended functionality, such as Jalview applets for checking query and template alignments, histogram views of alignments, and building 3D models with MODELLER.
In the CASP9 competition (Critical Assessment of Techniques for Protein Structure Prediction) in 2010, a fully automated version of HHpred based on HHsearch and HHblits was ranked best out of the 81 servers in template-based structure prediction, the category most relevant for biological applications, while having an average response time of minutes instead of days like most other servers .
Other popular programs for sensitive, iterative protein sequence searching are PSI-BLAST and HMMER. Since they are based on profile-to-sequence and HMM-to-sequence comparison, respectively, they have the advantage over HHblits and HHsearch of being able to search raw sequence databases.
The HH-suite source code for most Linux distributions and macOS, and its utility scripts written in Perl and Python can be found at:
https://github.com/soedinglab/hh-suite
In addition you have to install the Protein Data Bank parser for Python3 which can be found at:
https://github.com/soedinglab/pdbx
Databases can be downloaded at:
http://wwwuser.gwdg.de/~compbiol/data/hhsuite/databases/hhsuite_dbs/
HH-suite has been extensively tested on 64-bit Linux. We have done limited testing under BSD and macOS. HH-suite typically requires at least 4GB of main memory. Memory requirements can be estimated as follows:
Memory req.
= query_length * max_db_seq_length * num_threads * (4 (SSE2) or 8 (AVX2)) byte
+ query_length * max_db_seq_length * num_threads * 8 byte
+ 1 GB
Here, num_threads is the number of threads specified with option -cpu <int>.
Due to the high number of random file accesses by HHblits, it is important for good efficiency that the databases used by HHblits are placed in RAM disk or on a local solid state drive (SSD). See section Running HHblits efficiently on a computer cluster for details.
HHblits requires a CPU supporting at least the SSE2 (Streaming SIMD Extensions 2) instruction set. Every 64-bit CPU also supports SSE2. By default, the HH-suite binaries are compiled using the most recent supported instruction set on the computer used for compiling.
However, you may limit the used instruction set to SSE2 with the cmake
compile flag -DHAVE_SSE2=1. The precompiled standard binaries are compiled with this flag.
mkdir -p ~/programs/hh-suite && cd ~/programs/hh-suite
git clone https://github.com/soedinglab/hh-suite.git .
mkdir build && cd build
cmake ..
make
Set ${INSTALL_BASE_DIR} to the absolute path of the base directory
where you want install HH-suite. For example, to install into /usr/local/hhsuite:
INSTALL_BASE_DIR=/usr/local/hhsuite
cmake -DCMAKE_INSTALL_PREFIX="$INSTALL_BASE_DIR" ..
make install
The HH-suite binaries will then be placed in /usr/local/hhsuite/bin.
Some old HH-suite Python and Perl scripts will try to read the HHLIB environment variable to
locate HH-suite binaries and some required data files.
Set the HHLIB environment variable to ${INSTALL_BASE_DIR}, e.g, for bash use:
export HHLIB="${INSTALL_BASE_DIR}"
Put the location of your HH-suite binaries and scripts into your PATH:
export PATH="$PATH:${INSTALL_BASE_DIR}/bin:${INSTALL_BASE_DIR}/scripts"
To avoid typing these commands every time you open a new shell, you may
add the following lines to the .bashrc or
equivalent file in your home directory that is executed every time a
shell is started:
export HHLIB="/usr/local/hhsuite"
export PATH="$PATH:$HHLIB/bin:$HHLIB/scripts"
Since MPI is usually specific to the local cluster installation, the _mpi suffixed binaries have to be compiled from source. During the cmake invocation the build system will automatically check if MPI development files can be located and if found produce these binaries.
Generally you should refer to your local cluster admin team to find out how to load MPI development files for your local system. If you only want to use the _mpi suffixed binaries on a single node you can also try installing the development files as follows:
On Ubuntu/Debian run:
apt install mpi-default-bin mpi-default-dev
On macOS run:
brew install openmpi
The following HH-suite databases, which can be searched by HHblits and HHsearch, can be downloaded at:
http://wwwuser.gwdg.de/~compbiol/data/hhsuite/databases/hhsuite_dbs
-
Uniclust30: Based on UniProt KB, clustered to 30% seq. identity

- PDB70: Representatives from PDB (70% max. sequence identity), updated weekly
- scop70: Representatives from SCOP (70% max. sequence identity)
-
Pfam A: Protein Family database
Click on the ![]() links to go to the GitHub repository that contains the scripts to generate the respective database.
links to go to the GitHub repository that contains the scripts to generate the respective database.
The databases consist of usually to following six files, which all start with the name of the database, followed by different extensions:
<dbname>_cs219.ffdata packed file with column-state sequences for prefiltering
<dbname>_cs219.ffindex index file for packed column-state sequence file
<dbname>_a3m.ffdata packed file with MSAs in A3M format
<dbname>_a3m.ffindex index file for packed A3M file
<dbname>_hhm.ffdata packed file with HHM-formatted HMMs
<dbname>_hhm.ffindex index file for packed HHM file
The packed files <dbname>_cs219.ffdata, <dbname>_hhm.ffdata and
<dbname>_a3m.ffdata contain simply the concatenated A3M MSAs and HHMs,
respectively, with a \0 character at the end of each file. They
are therefore human-readable and are parsable for specific MSAs or
models using tools such as grep or search functions in text editors
(which however should be able to ignore the \0 character). The
.ffindex files contain indices to provide fast access to these packed
files.
To get started, download the Uniclust30 database. For example:
cd ~/programs/hh % change to HHsuite directory
mkdir databases; cd databases
wget http://wwwuser.gwdg.de/~compbiol/uniclust/2018_08/uniclust30_2018_08_hhsuite.tar.gz
tar xzvf uniclust30_2018_08_hhsuite.tar.gz
Note that, in order to generate multiple sequence alignments (MSAs) by iterative sequence searching using HHblits, you need to search the Uniclust30, since this databases covers essentially the largest part of the known sequence universe. The PDB70, PfamA, and SCOP are not appropriate to build MSAs by iterative searches.
Uniclust30 is obtained by clustering the UniProt Protein database. Clusters contain sequences that need to be almost full-length (80%) alignable and typically have pairwise sequence identities down to 30%. The clustering is done by MMseqs2, a very fast algorithm for all-against-all sequence comparison and clustering developed in our group. Sequences in each cluster are globally aligned into an MSA (using ClustalO). The clusters in Uniclust30 thus treat all member sequences equally. You need this type of database to build MSAs using iterative HHblits searches.
In the PDB70 and scop70 databases, each master sequence from the original sequence database is represented by an MSA. The MSAs are built by HHblits searches starting with the master sequence as a query. The MSAs and HMMs typically carry the name and annotation of the master sequence. In contrast to the clusters in the Uniclust30 database, sequences can in principle occur in several MSAs. These homologous sequences merely serve to contribute evolutionary information to the master sequence. As the sequences in this type of database do not cover the entire sequence space, they are not suited for iterative searches. See Building customized databases for how to build your own databases.
Note on efficiency: Always try to keep the HHsuite databases on a local SSD drive of the computer running HHblits/HHsearch, if possible. Otherwise file access times on your (shared) remote file server of local HD drive might limit the performance of HHblits/HHsearch.
When HHblits runs on many cores of a compute cluster and accesses the HHsuite database(s) via a central file system, hard disk random access times can quickly become limiting instead of the available compute power. To understand why, let us estimate the random file access bottleneck at an example. Suppose you want to search the Uniclust30 database with HHblits on four compute nodes with 32 CPU cores each. You let each job run on a single core, which is the most efficient way when processing many jobs. Each HHblits iteration through Uniclust30 on a modern 3GHz Intel/AMD core takes around, say, 60s on average. After each search iteration, HHblits needs to read around (3000) a3m or hhm files on disk that passed the prefilter. The total number of expected file accesses per second is therefore:
number of file accesses =
4 nodes * 32 CPU cores * 3000 files / 60s = 6400s
A typical server hard disk has a latency of around 5ms and can therefore randomly access only 200 files per second. Hence, if all cores need to access the files from the same RAID disk array, 4 CPU cores would already saturate the RAID’s disk drives! (You can slightly decrease latency by a factor of two by using RAID configurations with striping, but this won’t help much.)
To efficiently search with HHblits with millions of sequences in parallel on a compute cluster, we therefore recommend one of the first two options.
-
The ideal solution is to automatically load the needed HHsuite databases from the file system into the virtual RAM drive of each compute node upon boot-up. This requires enough memory in your computer. Software for RAM drives exist on all major operating systems. On Linux from kernel 2.4 upwards, a RAM drive is installed by default at
/dev/shm. When memory is low, this software can intelligently swap rarely used files to a hard drive. -
Another good option is to hold the HHsuite databases on local solid state drive (SSD) of each of the cluster’s compute nodes that can accept HHsuite jobs. The local SSDs also remove the file access bottleneck, as SSDs have typical random access times below 100ms. Reading 3000 files from a SSD after prefiltering thus should take below 0.3s.
-
Another good option might be to use a RAM disk (or SSD) on your file server to hold the HHsuite databases. Then file access time is not problematic anymore, but you need to estimate if data throughput might get limiting. The four servers from our previous example would need to read data at a rate:
transfer rate = 4 nodes * 32 CPU cores * 3000 files * 5kB (av. file size) / 60s
= 256MB/s
If you have 16 instead of 4 compute servers or if data transfer from the file server has to be shared with many other compute servers, the network transfer may be limiting the performance.
-
A not so good option is to keep the HHsuite databases on local hard disk drives (HDDs). In this way, you at least distribute the file accesses to as many HDDs as you have compute nodes. But as we have seen, already at around 4 cores per HDD, you will be limited by file access rather than CPU power.
-
You (or your sysadmin) should at least make sure that your compute nodes have sufficient memory reserves and proper cache settings in order to keep the prefiltering database file in cache (size: a few GB). You can check this by running HHblits on a fresh compute node twice and observing that the second run takes much less time to read the prefiltering database file. But this only solves the problem of having to read the prefiltering file every time hhblits is called. But, as discussed, the usually more serious bottleneck is reading the a3m and hhm files that passed the prefilter via the file system. These can also be cached, but since each time only a small subset of all files are read, caching is inefficient for these.
- hhblits: Iteratively) search an HHsuite database with a query sequence or MSA
- hhsearch: Search an HHsuite database with a query MSA or HMM
- hhmake: Build an HMM from an input MSA
- hhfilter: Filter an MSA by max sequence identity, coverage, and other criteria
- hhalign: Calculate pairwise alignments etc. for two HMMs/MSAs
- hhconsensus: Calculate the consensus sequence for an A3M/FASTA input file
- reformat.pl: Reformat one or many MSAs
- addss.pl: Add PSIPRED predicted secondary structure to an MSA or HHM file
- hhmakemodel.pl: Generate MSAs or coarse 3D models from HHsearch or HHblits results
- hhmakemodel.py: Generates coarse 3D models from HHsearch or HHblits results and modifies cif files such that they are compatible with MODELLER
- hhsuitedb.py: Build HHsuite database with prefiltering, packed MSA/HMM, and index files
- splitfasta.pl: Split a multiple-sequence FASTA file into multiple single-sequence files
- renumberpdb.pl: Generate PDB file with indices renumbered to match input sequence indices
- HHPaths.pm: Configuration file with paths to the PDB, BLAST, PSIPRED etc.
- mergeali.pl: Merge MSAs in A3M format according to an MSA of their seed sequences
- pdb2fasta.pl: Generate FASTA sequence file from SEQRES records of globbed pdb files
- cif2fasta.py: Generate a FASTA sequence from the pdbx_seq_one_letter_code entry of the entity_poly of globbed cif files
- pdbfilter.pl: Generate representative set of PDB/SCOP sequences from pdb2fasta.pl output
- pdbfilter.py: Generate representative set of PDB/SCOP sequences from cif2fasta.py output
Call a program without arguments or with -h option to get more detailed explanations.
We will use the MSA query.a3m in the data/ subdirectory of the
HHsuite as an example query. To search for sequences in the
scop70_1.75 database that are homologous to the query sequence or MSA
in query.a3m, type
hhsearch -cpu 4 -i data/query.a3m -d databases/scop70_1.75 -o data/query.hhr
The database scop70_1.75 can be obtained from our download server.
If the input file is an MSA or a single sequence, HHsearch calculates an
HMM from it and then aligns this query HMM to all HMMs in the
scop70_1.75 database using the Viterbi algorithm. After the search,
the most significant HMMs are realigned using the more accurate Maximum
Accuracy (MAC) algorithm. After the realignment
phase, the complete search results consisting of the summary hit list
and the pairwise query-template alignments are written to the output
file, data/query.hhr. The hhr result file format was designed to be
human readable and easily parsable.
The -cpu 4 option tells HHsearch to start four OpenMP threads for
searching and realignment. This will typically results in almost
fourfold faster execution on computers with four or more cores. Since
the management of the threads costs negligible overhead.
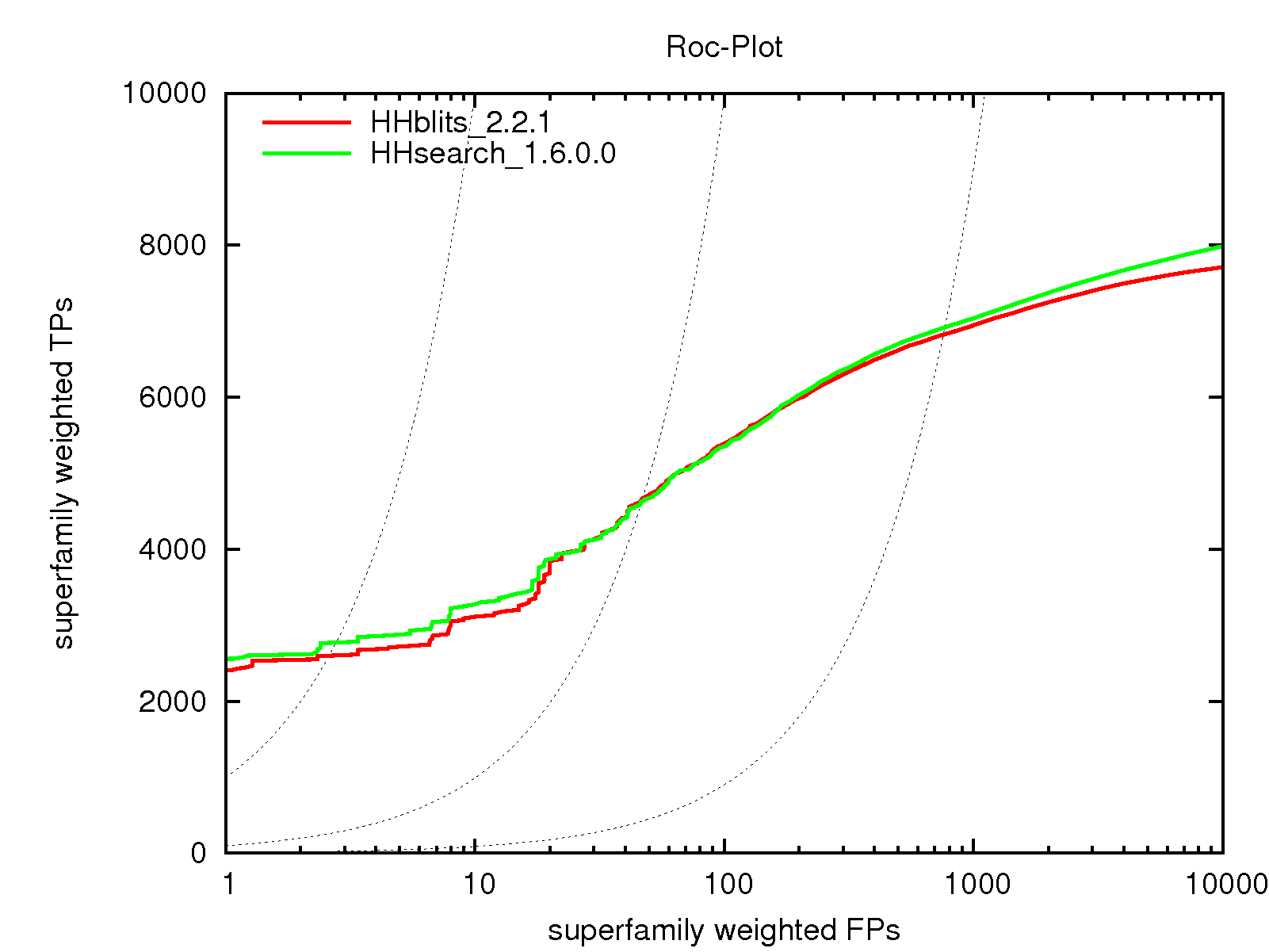
The HHblits tool can be used in much the same way as HHsearch. It takes the same input data and produces a results file in the same format as HHsearch. Most of the HHsearch options also work for HHblits, which has additional options associated with its extended functionality for iterative searches. Due to its fast prefilter, HHblits runs between 30 and 3000 times faster than HHsearch at the cost of only a few percent lower sensitivity.
The same search as above is performed here using HHblits instead of HHsearch:
hhblits -cpu 4 -i data/query.a3m -d databases/scop70_1.75 -o data/query.hhr -n 1
HHblits first scans the column state sequences in
scop70_1.75_cs219.ffdata with its fast prefilter. HMMs whose column
state sequences pass the prefilter are read from the packed file
scop70_1.75_hhm.ffdata (using the index file
scop70_1.75_hhm.ffindex) and are aligned to the query HMM generated
from query.a3m using the slow Viterbi HMM-HMM alignment algorithm. The
search results are written to the default output file query.hhr. The
option -n 1 tells HHblits to perform a single search iteration. (The
default is 2 iterations.)
To generate an MSA for a sequence or initial MSA in query.a3m, the
database to be searched should cover the entire sequence space, such as
the Uniclust30. The option -oa3m <msa_file> tells HHblits to
generate an output MSA from the significant hits, -n 1 specifies a
single search iteration.
hhblits -cpu 4 -i data/query.seq -d databases/uniclust30_2018_08/uniclust30_2018_08 -oa3m query.a3m -n 1
At the end of the search, HHblits reads from the packed database file
containing the MSAs the sequences belonging to HMMs with E-value below
the threshold. The E-value threshold for inclusion into the MSA can be
specified using the -e <E-value> option. After the search, query.a3m
will contain the MSA in A3M format.
We could do a second search iteration, starting with the MSA from the
previous search, to add more sequences. Since the MSA generated after
the previous search contains more information than the single sequence
in query.seq, searching with this MSA will probably result in many
more homologous database matches.
hhblits -cpu 4 -i query.a3m -d databases/uniclust30_2018_08/uniclust30_2018_08 -oa3m query.a3m -n 1
Instead, we could directly perform two search iterations starting from
query.seq:
hhblits -cpu 4 -i data/query.seq -d databases/uniclust30_2018_08/uniclust30_2018_08 -oa3m query.a3m -n 2
See Why do I get different results when I perform 2 and 1+1 iterations with HHblits? for an explanation why the results can be slightly different.
In practice, it is recommended to use between 1 and 4 iterations for building MSAs, depending on the trade-off between reliability and specificity on one side and sensitivity for remotely homologous sequences on the other side. The more search iterations are done, the higher will be the risk of non-homologous sequences or sequence segments entering the MSA and recruiting more of their kind in subsequent iterations. This is particularly problematic when searching with sequences containing short repeats, regions with amino acid compositional bias and, although less dramatic, with multiple domains. Fortunately, this problem is much less pronounced in hhblits as compared to PSI-BLAST due to hhblits’s lower number of iterations, its more robust Maximum Accuracy alignment algorithm, and the higher precision of its HMM-HMM alignments.
The parameter -mact (maximum accuracy threshold) lets you choose the
trade-off between sensitivity and precision. With a low mact-value
(e.g. -mact 0.01) very sensitive, but not so precise alignments are
generated, whereas a search with a high mact-value (e.g. -mact 0.9)
results in shorter but very precise alignments. The default value of
-mact in HHblits is 0.35.
To avoid unnecessarily large and diverse MSAs, HHblits stops iterating
when the diversity of the query MSA – measured as number of effective
sequences (N_eff) – grows past a threshold of 10.0. This threshold can be modified with the --neffmax <float>
option. See HHsearch/HHblits model format for a description of how the number of effective sequences is calculated in HH-suite.
To avoid that the final MSAs grows unnecessarily large, by default, the
database MSAs that are going to be merged with the query MSA into the
result MSA are filtered with the active filter options (by default
-id 90 and -diff 1000). The -all option turns off the filtering of
the result MSA. Use this option if you want to get all sequences in the
significantly similar Uniclust30 clusters:
hhblits -cpu 4 -i data/query.seq -d databases/uniclust30_2018_08/uniclust30_2018_08 -oa3m query.a3m -all
The A3M format uses small letters to mark inserts and capital letters to
designate match and delete columns (see Multiple sequence alignment formats),
allowing you to omit gaps aligned to insert columns. The A3M format
therefore uses much less space for large alignments than FASTA but looks
misaligned to the human eye. Use the reformat.pl script to reformat
query.a3m to other formats, e.g. for reformatting the MSA to Clustal
and FASTA format, type:
reformat.pl a3m clu query.a3m query.clu
reformat.pl a3m fas query.a3m query.fas
Next, to add secondary structure information to the MSA we call the
script addss.pl. For addss.pl to work, you have to make sure that
the paths to BLAST and PSIPRED in the file $HHLIB/scripts/HHPaths.pm
are correctly filled in:
addss.pl query.a3m
When the sequence has a SCOP or PDB identifier as first word in its
name, the script tries to add the DSSP states as well. Open the
query.a3m file and check out the two lines that have been added to the
MSA.
❗️ While secondary structure scoring does improve homology detection a bit, this step is extremely time consuming and currently hard to set up due to hardcoded old versions of tools. We recommend to skip this step.
Now you can generate a hidden Markov model (HMM) from this MSA:
hhmake -i query.a3m -o query.hhm
The default output file is query.hhm. By default, the option -M first will be used. This means that exactly those columns of the MSAs
which contain a residue in the query sequence will be assigned to Match
/ Delete states, the others will be assigned to Insert states. (The
query sequence is the first sequence not containing secondary structure
information.) Alternatively, you may want to apply the 50%-gap rule by
typing -M 50, which assigns only those columns to Insert states which
contain more than 50% gaps. The -M first option makes sense if your
alignment can best be viewed as a seed sequence plus aligned homologs to
reinforce it with evolutionary information. This is the case in the SCOP
and PDB versions of our HMM databases, since here MSAs are built around
a single seed sequence (the one with known structure). On the contrary,
when your alignment represents an entire family of homologs and no
sequence in particular, it is best to use the 50% gap rule. This is the
case for Pfam or SMART MSAs, for instance. Despite its simplicity, the
50% gap rule has been shown to perform well in practice.
When calling hhmake, you may also apply several filters, such as
maximum pairwise sequence identity (-id <int>), minimum sequence
identity with query sequence (-qid <int>), or minimum coverage with
query (-cov <int>). But beware of reducing the diversity of your MSAs
too much, as this will lower the sensitivity to detect remote homologs.
A three-dimensional (3D) structure greatly facilitates the functional characterization of proteins. However, for many proteins there are no experimental structures available, and thus, comparative modeling to known protein structures may provide useful insights. In this method, a 3D structure of a given protein sequence (target) is predicted based on alignments to one or more proteins of known structures (templates). In the following, we demonstrate how to create alignments for a unresolved protein with HHblits and the PDB70 database. We then convert search results from HHblits to build a comparative model using MODELLER (v9.16).
In 1999, Wu et all. reported the genomic sequence and evolutionary analysis of lactate dehydrogenase genes from Trichomonas vaginalis (TvLDH). Surprisingly, the corresponding protein sequence was most similar to the malate dehydrogenase (TvMDH) of the same organism implying TvLDH arose from TvMDH by convergent evolution. In the meantime, the structure of TvLDH has been resolved, however, for instructional purposes suppose that there is no 3D structure for the protein.
To get started we obtain the protein sequence of TvLDH from GeneBank
(accession number
AF060233.1). We first
copy-paste the protein sequence into a new file named query.seq. The
content of this file should somewhat look similar to this.
>TvLDH
MSEAAHVLITGAAGQIGYILSHWIASGELYGDRQVYLHLLDIPPAMNRLTALTMELEDCAFPHLAGFVATTDP
KAAFKDIDCAFLVASMPLKPGQVRADLISSNSVIFKNTGEYLSKWAKPSVKVLVIGNPDNTNCEIAMLHAKNL
KPENFSSLSMLDQNRAYYEVASKLGVDVKDVHDIIVWGNHGESMVADLTQATFTKEGKTQKVVDVLDHDYVFD
TFFKKIGHRAWDILEHRGFTSAASPTKAAIQHMKAWLFGTAPGEVLSMGIPVPEGNPYGIKPGVVFSFPCNVD
KEGKIHVVEGFKVNDWLREKLDFTEKDLFHEKEIALNHLAQ
The search results obtained by querying the TvLDH sequence against the PDB70 will be significantly better if we use a MSA instead of a single sequence. For this reason, we first query the protein sequence of TvLDH against the Uniclust30 database which covers the whole protein sequence space. By executing
hhblits -i query.seq -d databases/uniclust30_2018_08/uniclust30_2018_08 -oa3m query.a3m -cpu 4 -n 1
we obtain a MSA in a3m format which contains several sequences that are
similar to TvLDH. Now we can use the query.a3m and search the PDB70
database for similar protein structures.
hhblits -i query.a3m -o results.hhr -d databases/pdb70 -cpu 4 -n 1
Note that we now output a hhr file results.hhr instead of an a3m file.
Before we convert the search results to a format that is readable by
MODELLER, let us quickly inspect results.hhr.
Query TvLDH
Match_columns 333
No_of_seqs 2547 out of 8557
Neff 11.6151
Searched_HMMs 1566
Date Tue Aug 16 11:35:02 2016
Command hhblits -i query.a3m -o results.hhr -d pdb70 -cpu 32 -n 2
No Hit Prob E-value P-value Score SS Cols Query HMM Template HMM
1 7MDH_C MALATE DEHYDROGENASE; C 100.0 1.5E-39 1.3E-43 270.1 0.0 326 3-333 31-358 (375)
2 4UUL_A L-LACTATE DEHYDROGENASE 100.0 1.8E-35 1.6E-39 244.5 0.0 332 1-332 1-332 (341)
3 4UUP_A MALATE DEHYDROGENASE (E 100.0 2.2E-35 1.9E-39 243.8 0.0 332 1-332 1-332 (341)
4 4UUM_B L-LACTATE DEHYDROGENASE 100.0 5.7E-35 5.1E-39 241.5 0.0 333 1-333 1-333 (341)
5 1CIV_A NADP-MALATE DEHYDROGENA 100.0 1.6E-34 1.5E-38 241.3 0.0 326 2-332 40-367 (385)
6 1Y7T_A Malate dehydrogenase(E. 100.0 3.4E-34 3.1E-38 235.3 0.0 324 1-331 1-325 (327)
7 4I1I_A Malate dehydrogenase (E 100.0 3.8E-34 3.4E-38 236.4 0.0 319 2-327 22-343 (345)
8 1BMD_A MALATE DEHYDROGENASE (E 99.9 5.9E-34 5.3E-38 233.9 0.0 324 1-331 1-325 (327)
9 4H7P_B Malate dehydrogenase (E 99.9 9.9E-34 8.9E-38 233.9 0.0 319 2-327 22-343 (345)
10 2EWD_B lactate dehydrogenase, 99.9 1.1E-33 9.5E-38 231.1 0.0 306 1-328 1-314 (317)
(...)
We find that there are several templates that have a high similarity to our query. Interestingly, the hit with the most significant E-value score is also malate dehydrogenase. We will use this structure as a basis for our comparative model. In order to build the model we first have to obtain the template structure(s). We can get 7MDH by typing the following commands
mkdir templates
cd templates
wget http://files.rcsb.org/download/7MDH.cif
cd ..
To convert our search results results.hhr into an alignment that is
readable by MODELLER we use hhmakemodel.py.
python3 hhmakemodel.py results.hhr templates/ TvLDH.pir ./ -m 1
This script takes four positional arguments: the results file in hhr
format, the path to the folder containing all templates in cif format,
the output pir file, and folder where the processed cif files should be
written to. The -m flag tells hhmakemodel.py to only include the
first hit in the pir alignment. The pir file together with processed
cifs can be used as an input for MODELLER (please refer to the MODELLER
documentation for further help).
Iterative search methods such as PSI-BLAST and HHblits may generate alignments containing non-homologous sequence stretches or even large fractions of non-homologous sequences. The cause for this is almost always the overextension of homologous alignments into non-homologous regions. This has been termed homologous overextension in . This effect occurs particularly in multidomain and repeat proteins. A single overextended alignment in the search results leads to more of such sequences to be included in the profile of the next iteration – usually all homologous to the first problematic sequence but with even longer non-homologous stretches. Thus, after three or more iterations, large sections of the resulting MSA may be non-homologous to the original query sequence. This risk of homologous overextension is greatly reduced in HHblits in comparison to PSI-BLAST, because fewer iterations are usually necessary for HHblits and because HHblits uses the Maximum Accuracy alignment algorithm (see What does the maximum accuracy alignment algorithm do?), which is much less prone to overextend alignments than the Smith-Waterman/Viterbi algorithm used by most other programs including PSI-BLAST. Still, in important cases it is worth to visually check the MSAs of the query and the matched database protein for the presence of corrupted regions containing non-homologous sequence stretches.
The recommended procedure to visually check an MSA is the following. We first reduce the MSA to a small set of sequences that could still fit into a single window of an alignment viewer:
hhfilter -i query.a3m -o query.fil.a3m -diff 30
The option -diff causes hhfilter to select a representative set of at
least 30 sequences that best represent the full diversity of the MSA.
Sequences that contain non-homologous stretches are therefore usually
retained, as they tend to be the most dissimilar to the main sequence
cluster.
Next, we remove all inserts (option -r) with respect to the first,
master sequence in the MSA. The resulting MSA is sometimes called a
master-slave alignment:
reformat.pl -r query.fil.a3m query.fil.fas
This alignment is now very easy to inspect for problematic regions in
any viewer that allows to color the residues according to their
physico-chemical properties. We can recommend alnedit
or jalview, for example:
java -jar alnedit.jar query.fil.fas .
It is possible to build custom HH-suite databases using the same tools we use to build the standard HH-suite databases (except Uniclust30). An example application is to search for homologs among all proteins of an organism. To build your own HH-suite database from a set of sequences, you first need to generate an MSA with predicted secondary structure for every sequence in the set.
🗒 The following calls assume you have compiled the HH-suite with MPI support. You can substitute hhblits_mpi with hhblits_omp, ffindex_apply_mpi with ffindex_apply and cstranslate_mpi with cstranslate -f to use the OpenMP (single compute node) parallelization instead.
First of all we convert the input FASTA file to a FFindex database with
ffindex_from_fasta:
ffindex_from_fasta -s <db>_fas.ff{data,index} <db.fas>
Now, to build an MSA with HHblits for each sequence in <db>_fas.ff{data,index}, run
mpirun -np <number_threads> \
hhblits_mpi -i <db>_fas -d <path_to/uniclust30> -oa3m <db>_a3m_wo_ss -n 2 -cpu 1 -v 0
The MSAs are written to the ffindex <db>_a3m_wo_ss.ff{data,index}. To
be sure that everything went smoothly, check that the number of lines in
<db>_a3m.ffindex is the same as the number of lines in <db>_fas.ffindex.
The number of HHblits search iterations and the HMM inclusion E-value
threshold for HHblits can be changed from their default values (2 and
0.01, respectively) using the -n <int> and -e <float> options. A
higher number of iterations such as -n 3 will result in very high
sensitivity to discover remotely homologous relationships, but the
ranking among homologous proteins will often not reflect their degree of
relationship to the query. The reason is that any similarities that are
higher than the similarities among the sequences in the query and
database MSAs cannot be resolved.
If you already have MSAs in aligned FASTA format, place all of them first in a single folder that does not contain any other files to create a single FFindex database:
cd msa/
ffindex_build -s ../<db>_msa.ff{data,index} .
cd ..
Now you can use hhconsensus to add a consensus sequence and do the format conversion to the more space efficient A3M format:
OMP_NUM_THREADS=1 mpirun -np <number_threads> ffindex_apply_mpi <db>_msa.ff{data,index} \
-i <db>_a3m_wo_ss.ffindex -d <db>_a3m_wo_ss.ffdata \
-- hhconsensus -M 50 -maxres 65535 -i stdin -oa3m stdout -v 0
rm <db>_msa.ff{data,index}
Now, add PSIPRED-predicted secondary structure and if possible DSSP secondary structure annotation to all MSAs:
❗️ While secondary structure scoring does improve homology detection a bit, this step is extremely time consuming and currently hard to set up due to hardcoded old versions of tools. We recommend to skip this step. Rename _a3m_wo_ss.ff{data,index} to _a3m.ff{data,index} instead to proceed with the next step.
mpirun -np <number_threads> ffindex_apply_mpi <db>_a3m_wo_ss.ff{data,index} \
-i <db>_a3m.ffindex -d <db>_a3m.ffdata -- addss.pl -v 0 stdin stdout
rm <db>_a3m_wo_ss.ff{data,index}
Computing hidden markov models
We also need to generate an HHM model for each MSA:
mpirun -np <number_threads> ffindex_apply_mpi <db>_a3m.ff{data,index} \\
-i <db>_hhm.ffindex -d <db>_hhm.ffdata -- hhmake -i stdin -o stdout -v 0
In order to build the FFindex database containing the column state sequences for prefiltering for each a3m we run:
mpirun -np <number_threads> cstranslate_mpi -f -x 0.3 -c 4 -I a3m -i <db>_a3m -o <db>_cs219
❗️ Be careful with the parameters. Leaving anything out, might result in a database that looks superficially correct, but will perform very badly.
❗️ Before version 3.2 you would need to specify the -b parameter to cstranslate to produce a HHblits3 compatible database, without it you would produce a database that could be still post-processed to be readable by HHblits2.x. We have since dropped support for HHblits2.x and the -b parameter.
Next we want to reorder the entries in the hmm and a3m databases. For this purpose we
sort the files in the data files according to the number of columns in
the MSAs (i.e. the sequence length of the consensus sequence). The number of columns can be retrieved in the third row of <db>_cs219.ffindex.
sort -k3 -n -r <db>_cs219.ffindex | cut -f1 > sorting.dat
ffindex_order sorting.dat <db>_hhm.ff{data,index} <db>_hhm_ordered.ff{data,index}
mv <db>_hhm_ordered.ffindex <db>_hhm.ffindex
mv <db>_hhm_ordered.ffdata <db>_hhm.ffdata
ffindex_order sorting.dat <db>_a3m.ff{data,index} <db>_a3m_ordered.ff{data,index}
mv <db>_a3m_ordered.ffindex <db>_a3m.ffindex
mv <db>_a3m_ordered.ffdata <db>_a3m.ffdata
To make efficient sequence searches in the PDB we provide a precompiled PDB70 database containing PDB sequences clustered at 70% sequence identity. However, to find a larger variety of PDB templates, larger databases with more redundancy might be required. In this tutorial, we outline the steps required to build a custom PDB database. See the PDB70 GitHub repository for the actual scripts used to build this database.
First, download the entire PDB database from
RSCB in
cif file format (this is the successor of the pdb file format) by
executing
rsync --progress -rlpt -v -z --port=33444 rsync.wwpdb.org::ftp/data/structures/divided/mmCIF <cif_dir>
and unzip the files into a single directory <all_cifs>. Then, run
cif2fasta.py by typing
python3 cif2fasta.py -i <all_cifs> -o pdb100.fas -c <num_cores> -p pdb_filter.dat
The script scans the folder <all_cifs> for files with the suffix
*.cif and write each sequence and its associated chain identifier
annotated in pdbx_seq_one_letter_code entry from the entity_poly
table to the fasta file pdb100.fas. By specifying the optional -p
flag, cif2fasta.py creates an additional file pdb_filter.dat which
is required by pdbfilter.py in a later step. Note that cif2fasta.py
by default removes sequences which are shorter than 30 residues and/or
comprise only the residue ’X’.
If you wish to exhaustively search the PDB, skip the following steps and
continue with the instructions described in section
Building customized databases. However, to increase the speed of database
searches, e.g. on systems with limited resources, you can reduce the
number of sequences by clustering them with MMseqs2 and selecting
representative sequences with pdbfilter.py. To cluster the sequences
of pdb100.fas at a sequence identity of X and a coverage of Y run
MMseqs2 using these options.
mmseqs createdb pdb100.fas <clu_dir>/pdb100
mmseqs cluster <clu_dir>/pdb100 <clu_dir>/pdbXX_clu /tmp/clustering -c Y --min-seq-id X
mmseqs createtsv <clu_dir>/pdb100 <clu_dir>/pdb100 <clu_dir>/pdbXX_clu <clu_dir>/pdbXX_clu.tsv
MMseqs2 yields a tab separated file pdbXX_clu.tsv which contains cluster
assignments for all sequences. Representative sequences are selected by
pdbfilter.py which chooses up to three sequences for each cluster by
identifying the ones having either the highest resolution
A, the largest R-free value or the largest "completeness".
We compute the completeness of a protein structure by dividing the number of residues that are found in the ATOM section by the total number of residues declared in the pdbx_seq_one_letter_code entry of the entity_poly table.
python3 pdbfilter.py pdb100.fas pdbXX_clu.tsv pdb_filter.dat pdbXX.fas -i pdb70_to_include.dat -r pdb70_to_remove.dat
pdbfilter.py takes the original fasta file (pdb100.fas) and the
annotation file pdb_filter.dat which both were created by
cif2fasta.py, and the cluster assignments from MMseqs2
(pdb70_clu.tsv) as input and outputs the final pdbXX.fas. Use this
fasta file to complete the creation of your database (see section
Building customized databases).
Assume you have a list of filenames in files.dat you want to remove
from an HHsuite database.
ffindex_modify -s -u -f files.dat <db>_a3m.ffindex
ffindex_modify -s -u -f files.dat <db>_hhm.ffindex
ffindex_modify -s -u -f files.dat <db>_cs219.ffindex
This deletes the file entries from the ffindex files, however the files are still in the ffdata file. This way HHblits and HHsuite won’t be able to use them. If you want to get rid of them in the ffdata file you may rebuild the databases.
ffindex_build -as <db>_cs219_ordered.ff{data,index} -i <db>_cs219.ffindex -d <db>_cs219.ffdata
mv <db>_c219_ordered.ffindex <db>_cs219.ffindex
mv <db>_cs219_ordered.ffdata <db>_cs219.ffdata
sort -k3 -n <db>_cs219.ffindex | cut -f1 > sorting.dat
ffindex_order sorting.dat <db>_hhm.ff{data,index} <db>_hhm_ordered.ff{data,index}
mv <db>_hhm_ordered.ffindex <db>_hhm.ffindex
mv <db>_hhm_ordered.ffdata <db>_hhm.ffdata
ffindex_order sorting.dat <db>_a3m.ff{data,index} <db>_a3m_ordered.ff{data,index}
mv <db>_a3m_ordered.ffindex <db>_a3m.ffindex
mv <db>_a3m_ordered.ffdata <db>_a3m.ffdata
If you want to check your HHsuite database you may run:
hhsuitedb.py -o <db> --cpu 1
hhsuitedb.py supports the flag --force that tries to fix the
database. This may lead to the deletion of MSAs from the database.
Additionally you may add a3m, hhm and cs219 files with globular expressions to an existing database. Please keep in mind that related a3m, hhm and cs219 files should have the same name. To make it easier you may add just a3m files and the corresponding hhm and cs219 files will be calculated by the script.
hhsuitedb.py -o <db> --cpu 1 --ia3m=<a3m_glob> --ics219=<cs219_glob> --ihhm=<hhm_glob>
If you found a bug, please report it to us using GitHub issues.
When searching for remote homologs, it is wise to make use of as much information about the query and database proteins as possible in order to better distinguish true from false positives and to produce optimal alignments. This is the reason why sequence-sequence comparison is inferior to profile-sequence comparison. Sequence profiles contain for each column of a multiple alignment the frequencies of the 20 amino acids. They therefore contain detailed information about the conservation of each residue position, i.e., how important each position is for defining other members of the protein family, and about the preferred amino acids. Profile Hidden Markov Models (HMMs) are similar to simple sequence profiles, but in addition to the amino acid frequencies in the columns of a multiple sequence alignment they contain information about the frequency of inserts and deletions at each column. Using profile HMMs in place of simple sequence profiles should therefore further improve sensitivity. Using HMMs both on the query and the database side greatly enhances the sensitivity/selectivity and alignment quality over sequence-profile based methods such as PSI-BLAST. HHsearch is the first software to employ HMM-HMM comparison and HHblits is the first profile-profile comparison method that is fast enough to do iterative searches to build MSAs.
Sequence search methods such as BLAST, FASTA, or PSI-BLAST are of prime importance for biological research because functional information of a protein or gene can be inferred from homologous proteins or genes identified in a sequence search. But quite often no significant relationship to a protein of known function can be established. This is certainly the case for the most interesting group of proteins, those for which no ortholog has yet been studied. In cases where conventional sequence search methods fail, HHblits and HHsearch quite often allow to make inferences from more remotely homologous relationships. HHblits builds better MSAs, with which more remote homologs can then be found using HHsearch or HHblits, e.g. by searching the PDB or domain databases such as Pfam. If the relationship is so remote that no common function can be assumed, one can often still derive hypotheses about possible mechanisms, active site positions and residues, or the class of substrate bound. When a homologous protein with known structure can be identified, its stucture can be used as a template to model the 3D structure of the protein of interest , since even protein domains that shared a common ancestor some 3 billion years ago mostly have similar 3D structures. The 3D model may then help to generate hypotheses to guide experiments.
Two protein sequences are homologous to each other if they descended from a common ancestor sequence. Generally, homologous proteins (or protein fragments) have similar structure because structures diverge much more slowly than their sequences . Depending on the degree of divergence between the sequences, the proteins may also have similar cellular functions, ligands, protein interaction partners, or enzymatic mechanisms. On the contrary, proteins that have a similar structure by convergence (i.e., by chance) are said to be analogous. They don’t generally share similar functions or biochemical mechanisms and are therefore much less helpful for making inferences. HHsearch and HHblits are tools for homology detection and as such do not normally detect analogous relationships.
Here is a list of things to check if a database match really is at least locally homologous.
Check probability and E-value: HHsearch and HHblits can detect homologous relationships far beyond the twilight zone, i.e., below 20% sequence identity. Sequence identity is therefore not an appropriate measure of relatedness anymore. The estimated probability of the template to be (at least partly) homologous to your query sequence is the most important criterion to decide whether a template HMM is actually homologous or just a high-scoring chance hit. When it is larger than 95%, say, the homology is nearly certain. Roughly speaking, one should give a hit serious consideration (i.e., check the other points in this list) whenever (1) the hit has (>50%) probability, or (2) it has (>30%) probability and is among the top three hits. The E-value is an alternative measure of statistical significance. It tells you how many chance hits with a score better than this would be expected if the database contained only hits unrelated to the query. At E-values below one, matches start to get marginally significant. Contrary to the probability, when calculating the E-value HHsearch and HHblits do not take into account the secondary structure similarity. Therefore, the probability is a more sensitive measure than the E-value.
Check if homology is biologically suggestive or at least reasonable: Does the database hit have a function you would expect also for your query? Does it come from an organism that is likely to contain a homolog of your query protein?
Check secondary structure similarity: If the secondary structure of query and template is very different or you can’t see how they could fit together in 3D, then this is a reason to distrust the hit. Note however that if the query alignment contains only a single sequence, the secondary structure prediction is quite unreliable and confidence values are overestimated.
Check relationship among top hits: If several of the top hits are homologous to each other, (e.g. when they are members of the same SCOP superfamily), then this will considerably reduce the chances of all of them being chance hits, especially if these related hits are themselves not very similar to each other. Searching the SCOP database is very useful precisely for this reason, since the SCOP family identifier (e.g. a.118.8.2) allows to tell immediately if two templates are likely homologs.
Check for possible conserved motifs: Most homologous
pairs of alignments will have at least one (semi-)conserved motif in
common. You can identify such putative (semi-)conserved motifs by the
agglomeration of three or more well-matching columns (marked with a
| sign between the aligned HMMs) occurring within a few
residues, as well as by matching consensus sequences. Some false
positive hits have decent scores due to a similar amino acid composition
of the template. In these cases, the alignments tend to be long and to
lack conserved motifs.
Check residues and role of conserved motifs: If you can identify possible conserved motifs, are the corresponding conserved template residues involved in binding or enzymatic function?
Check query and template alignments: A corrupted query or template alignment is the main source of high-scoring false positives. The two most common sources of corruption in an alignment are (1) non-homologous sequences, especially repetitive or low-complexity sequences in the alignment, and (2) non-homologous fragments at the ends of the aligned database sequences. Check the query and template MSAs in an alignment viewer such as Jalview.
Realign with other parameters: change the alignment parameters. Choose global instead of local mode, for instance, if you expect your query to be globally homologous to the putative homolog. Try to improve the probability by changing the values for minimum coverage or minimum sequence identity. You can also run the query HMM against other databases.
Build the query and/or database MSAs more aggressively: If your query (or template) MSA is not diverse enough, you could increase sensitivity substantially by trying to include more remotely homologous sequences into the MSA. Have non-homologous sequences or sequence segments been accidentally included? You can also try to build a more diverse MSA manually: Inspect the HHblits results after the first iteration and consider including hits above the E-value inclusion threshold of 0.001, based on biological plausibility, relatedness of the organism, a reasonable looking alignment, or just guessing. Then start the second HHblits search iteration HHblits with this manually enriched alignment.
Verify predictions experimentally: The ultimate confirmation of a homologous relationship or structural model is, of course, the experimental verification of some of its key predictions, such as validating the binding to certain ligands by binding assays, measuring biochemical activity, or comparing the knock-out phenotype with the one obtained when the putative functional residues are mutated.
HHblits and HHsearch use a better alignment algorithm than the quick and
standard Viterbi method to generate the final HMM-HMM alignments. Both
realign all displayed alignments in a second stage using the more
accurate Maximum Accuracy (MAC) algorithm . The Viterbi algorithm is
employed for searching and ranking the matches. The realignment step is
parallelized (-cpu <int>) and typically takes a few seconds only.
Please note: Using different alignment algorithms for scoring and
aligning has the disadvantage that the pairwise alignments that are
displayed are not always very similar to those that are used to
calculate the scores. This can lead to confusing results where
alignments of only one or a few residues length may have obtained
significant E-values. In such cases, run the search again with the
-norealign option, which will skip the MAC-realignment step. This will
allow you to check if the Viterbi alignments are valid at all, which
they will probably not be. The length of the MAC alignments can
therefore give you additional information to decide if a match is valid.
In order to avoid confusion for users of our HHpred server , the
-norealign option is the default there, whereas for you pros who dare
to use the command line package, realigning is done by default.
The posterior probability threshold is controlled with the -mact
option. This parameter controls the alignment algorithm’s greediness.
More precisely, the MAC algorithm finds the alignment that maximizes the
sum of posterior probabilities minus mact for each aligned pair. Global
alignments are generated with -mact 0, whereas -mact 0.5 will produce
quite conservative local alignments.
The -global and -local options now refer to both the Viterbi search stage as well as the MAC realignment stage. With -global (-local), the posterior probability matrix will be calculated for global (local) alignment. When -global is used in conjunction with -realign, the mact parameter is automatically set to 0 in order to produce global alignments. In other words, both following two commands will give global alignments:
hhsearch -i <query> -d <db> -realign -mact 0
hhsearch -i <query> -d <db> -realign -global
The first version uses local Viterbi to search and then uses MAC to realign the proteins globally (since mact is 0) on a local posterior probability matrix. The second version uses global Viterbi to search and then realigns globally (since mact is automatically set to 0) on a global posterior matrix. To detect and align remote homologs, for which sometimes only parts of the sequence are conserved, the first version is clearly better. It is also more robust. If you expect to find globally alignable sequence homologs, the second option might be preferable. In that case, it is recommended to run both versions and compare the results.
The number of effective sequences of the full alignment, which appears
as NEFF in the header of each hhm file, is the average of local values
Neff_M(i) over all alignment positions i. The values Neff_M(i)
are given in the main model section of the hhm model files (subsection
HHsearch/HHblits model format). They quantify the local diversity of the alignment
in a region around position i. More precisely, Neff_M(i) measures
the diversity of subalignment Ali_M(i) that contains all sequences
that have a residue at column i of the full alignment. The
subalignment contains all columns for which at least 90% of these
sequences have no end gap. End gaps are gaps to the left of the first
residue or to the right of the last residue. The latter condition
ensures that the sequences in the subalignment Ali_M(i) cover most
of the columns in it. The number of effective sequences in the
subalignment Ali_M(i) is exp of the average sequence entropy over
all columns of the subalignment. Hence, Neff_M is bounded by 0 from
below and 20 from above. In practice, it is bounded by the entropy of a
column with background amino acid distribution f_a:
N_eff < sum_(a=1)^20 f_a*log(f_a) ≈ 16
Similarly, Neff_I(i) gives the diversity of the subalignment Ali_I(i) of all
sequences that have an insert at position i, and Neff_D(i) refers
to the diversity of subaligment Ali_D(i) of all sequences that have
a Delete (a gap) at position i of the full alignment.
HHblits and HHsearch have been designed to work with multi-domain queries. However, the chances for false positives entering the query alginment during the HHblits iterations is greater for multi-domain proteins. For long sequences, it may therefore be of advantage to first search the PDB or the SCOP domain database and then to cut the query sequence into smaller parts on the basis of the identified structural domains. Pfam or CDD are - in our opinion - less suitable to determine domain boundaries.
Despite the biased amino acid composition in their integral membrane helices, membrane proteins behave very well in HMM-HMM comparisons. In many benchmark tests and evolutionary studies we have not noticed any higher rates of false positives than with cytosolic proteins.
For example when domain A is predicted from
residues 2-50 with 98% probability and domain B from 2-200 with 95%
probability? The probability that a pair of residues is correctly
aligned is the product of the probability for the database match to be
homologous (given by the values in the Probab column of the hit list)
times the posterior probability of the residue pair to be correctly
aligned given the database match is correct in the first place. The
posterior probabilities are specified by the confidence numbers in the
last line of the alignment blocks (0 corresponds approximately to 0-10%,
9 to 90-100%). Therefore, an obvious solution would be to prune the
alignments in the overlapping region such that the sum of total
probabilities is maximized. There is no script yet that does this
automatically.
I would use a similarity measure like the raw score per alignment length. You might also add the secondary structure score to the raw score with some weight. Whereas probabilities, E-values, and P-values are useful for deciding whether a match is a reliable homolog or not, they are not suitable for measuring similarities because they strongly depend on the length of the alignment, roughly like:
P-value ∝ exp^(λ * average_similarity * length),
with some constant lambda of order 1. The probability has an even
more complex dependence on length. Also, the “Similarity” given above
the alignment blocks is a simple substitution matrix score per column
between the query and template master sequences and does not capture the
evolutionary information contained in the MSAs (see
HMM-HMM pairwise alignments). One note of caution: Large, diverse MSAs are
usually more sensitive to find homologs than narrower ones. Therefore, I
would limit the diversity of all HMMs to some reasonable number (perhaps
around 5 or 7, depending on how far diverged your HMMs are). This
filtering can be done using:
hhfilter -i <MSA.a3m> -o <MSA_filt.a3m> -neff 5
For example to select templates for homology
modeling, it could make sense to weight residues around the known
functional site more strongly. This might also positively impact the
query-template alignment itself. Well, directly weighting query or
template match states is not possible. What you can do with a bit of
file fiddling, however, may even be better: You can modify the amount of
pseudocounts that hhblits, hhsearch, or hhalign will automatically add
to the various match state columns in the query or template profile HMM.
This is equivalent to assuming a higher conservation for these
positions, since pseudocounts are basically mutations that are added to
the observed counts in the training sequences to generalize to more
remotely related sequences. In HHsuite, the more diverse the sequence
neighborhood around a match state column i is, as measured by the
number of effective sequences Neff_M(i) (see How is the MSA diversity Neff calculated?),
the fewer pseudocounts are added. By manually increasing Neff_M(i) for
a match state column you increase the assumed conservation of this
position in the protein. The Neff_M(i) values are found in the
hhm-formatted HMM model files in the eighth column of the second line of
each match state block in units of 0.001 (see HHsearch/HHblits model format).
You could, for example, simply increase Neff_M(i)
values by a factor of two for important, conserved positions. Setting
Neff_M(i) to 99999 will reduce pseudocounts to effectively zero. A
word of caution is advised: HMMs built from a diverse set of training
sequences already contain quantitative information about the degree of
conservation for each position.
No. If you don’t specify otherwise, the two parameters
of the extreme-value distribution for the query are estimated by a
neural network from the lengths and diversities N_eff of
query and database HMMs that was trained on a large set of example
queries-template pairs, in an approach similar to the one used in .
However, the old calibration is still available as an option in
HHsearch.
Should I use the -global option to build MSAs if I am intersted in global alignments to these database HMMs?
Never use -global when building MSAs by iterative
searches with HHblits. Remember that global alignments are very greedy
alignments, where alignments stretch to the ends of either query of
database HMM, no matter what. Global alignments will therefore often
lead to non-homologous segments getting included in the MSA, which is
catastrophic, as these false segments will lead to many false positive
matches when searching with this MSA. But you may use -global when
searching (with a single iteration) for global matches of your query
HMMs through your customized HHsuite database, for example.
One iteration! Doing more than one search iterations in general only makes sense in order to build up a diverse MSA of homologous sequences by searching through the Uniclust30 or a clustered NR databases.
For reasons of efficiency, the MSA A3M and HHM model
files are packed into two files, <db>_a3m.ffdata and
<db>_hhm.ffdata, each with its own index file. The contained files are
easy to extract from the packed files. For example, to dump
d1aa7a_.a3m in scop70 to standard output, type
ffindex_get scop70_a3m.ffdata scop70_a3m.ffindex d1aa7a_.a3m
You may write the extracted file to a separate file by appending '> d1aa7a_.a3m' to the above command. If you want to extract all files in
the ffindex data structures to directory tmp/scop70, type
ffindex_unpack scop70_a3m.ffdata scop70_a3m.ffindex tmp/scop70/ .
The procedure to cluster the UniProt databases is more complicated than building a database for a genome or for the sequences in the PDB. As its first step it involves clustering these huge databases down to 20%-30% sequence identity. The clustering is done in our lab using MMseqs2. We provide this database - the Uniclust30 - including all scripts to build it. These scripts generate A3M files, HHM files, and consensus sequences. Because of the large number of files to generate, these scripts need to be run on a computer cluster and this would require considerable computer savviness.
Currently we do not provide environmental databases for the HH-suite. Please use MMseqs2 to search through such datasets. MMseqs2 can easily search through the largest data such, such as assembled proteins from environmental metagenomes. We offer a Soil Reference Catalog (SRC) and a Marine Eukaryotic Reference Catalog, with billions of environmental proteins.
The pdb70 database is clustered to 70% maximum
pairwise sequence identity to reduce redundancy. The PDB identifiers at
the end of the description line of sequence X list the sequences
with lower experimental resolution that were removed for having higher
than 70% sequence identity with PDB sequence X. The asterisks
indicate that a multi-atom ligand is bound in the structure.
When taking A as the query, the pairwise alignment and significance scores with template B in the database can be different than when B is the query and A the template. By default hhblits, hhsearch, and hhalign add context-specific pseudocounts to the query MSA or HMM whereas the much faster substitution matrix pseudocounts are added to the database HMMs/MSAs for reasons of speed. If you want to combine the significance estimates from the forward and reverse comparisons, we recommend to take the geometric mean of Probability, P-value and E-value. Since the less diverse HMM or MSA will profit more from the context-specific pseudocounts than the more diverse HMM or MSA, it might be even better to take the estimate from the comparison in which the HMM/MSA with the smaller diversity (measured as number of effective sequences, NEFF) is used as the query.
Each line in the summary hit list refers to an alignment with an HMM in the database, not to the database HMM itself. Sometimes, alternative (suboptimal) alignments covering a different part of either the query or the database HMM may appear in the hit list. Usually, both the optimal and the alternative alignments are correct, in particular when the query or the database HMMs represent repeat proteins.
Why do I get different results using HHblits and HHsearch when I search with the same query through the same database?
There are two reasons. First, some hits that hhsearch shows
might not have passed the prefilter in hhblits. The option
-prepre_smax_thresh <bits> lets you modify the minimum score threshold
of the first, gapless alignment prefilter (default is 10 bits). Option
-pre_evalue_thresh <E-value> sets the maximum E-value threshold for
the second, gapped alignment prefilter (default is 1000). Second, while
the probabilities and P-values of hhblits and hhsearch should be the
identical for the same matches, the E-values are only similar. The
reason is that hhblits heuristically combines the E-values of the second
prefilter with the E-values from the full HMM-HMM Viterbi alignment into
total E-values . These E-values are slightly better in distinguishing
true from false hits, because they combine the partly independent
information from two comparisons.
When you do a single iteration and start a second
iteration with the A3M output file from the first iteration, the
alignments should be nearly the same as if you do two iterations
(-n 2) Why not exactly the same? Because when doing multiple
iterations in a single hhblits run, alignments for each hit are only
calculated once. When the same database HMM is found in later search
iterations, the “frozen”, first alignment is used instead of recomputing
it. This increases robustness with respect to homologous overextension.
However, the significance values can be slightly different between the
cases with 2 and 1+1 iterations. The reason is that the HHblits
P-value is calculated by a weighted combination of the P-value of the
Viterbi pairwise HMM alignment and the prefilter E-value. The prefilter
E-values differ in the two cases described, since in the first case the
E-value is calculated for the first prefilter search (before the first
full search) and in the second case it is calculated for the second
prefilter search, performed with the query MSA obtained after the
first search iteration.
Often, segmentation faults can be caused by too little available memory. HHblits needs memory to read the entire column state file into memory for prefiltering. For Viterbi HMM-HMM alignment it needs about:
Memory Viterbi = Query_length * max_db_seq_length * (num_threads+1) * 5 byte
Here, num_threads is the number of threads specified with option -cpu <int>. For realignment, HHblits needs in addition:
Memory Realign = Query_length * max_db_seq_length * (num_threads+1) * 8 byte
But the amount of memory for realignment can be limited using the
-maxmem <GB> option, which will cause the realignment to skip the
longest sequences.
Make sure to enable the memory overcommitment on your machine. On Linux either add vm.overcommit_memory=1 to /etc/sysctl.conf and then reboot or run the command sysctl vm.overcommit_memory=1.
Under Linux, you can check your memory limit settings
using ulimit -a. Make sure you have at least 4GB per thread. Try
ulimit -m 4000000 -v 4000000 -s 8192 -f 4000000
Also try using -cpu 1 -maxmem 1 to reduce memory requirements. If it
still does not work, follow the tipps in subsection
How do I report a bug? to report your problems to us.
I obtain different results for the same query sequence on the web server and on my local version using HHblits and HHsearch.
-
Check the exact command line calls for HHblits and HHsearch from the log file accessible on the results page of the server. If any of the commands differ, rerun your command line version with the same options.
-
Check if the query multiple sequence alignments on the server and in your versions contain the same number of sequences and the same diversity. If not, try to find out why (see previous point, for example.)
-
For some of the alignments with differing scores, check if the db aligment on the server is the same as the one in your local database. You can download the db MSA by clicking the alignment logo above the query-template pairwise profile-profile alignment.
I find an alignment of my query and a template with the exact same sequence, but they are aligned incorrectly.
I find a gap inserted at residue position 92 in query and position 99 in
template:
Q ss_pred CcEEEeeCCCC-CCCeEEEEEECCeeEEEEEEECCCCcEEEEEEeeCCHHHHHHHHhhCCCccceeEEEecccCCCCCCe
Q Pr_1 81 GAFLIRESESA-PGDFSLSVKFGNDVQHFKVLRDGAGKYFLWVVKFNSLNELVDYHRSTSVSRNQQIFLRDIEQVPQQPT 159 (217)
Q Consensus 81 ~~~~~~~~~~~-~~~~~~s~~~~~~~~~~~i~~~~~~~~~~~~~~f~s~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ 159 (217)
|.+++|.+... ++.+.+ +..++.++|+++.....+.|+.+...|.++..++.++...+....................
T Consensus 81 G~flvR~s~~~~~~~~~l-v~~~~~v~h~~i~~~~~g~~~~~~~~f~sl~~Lv~~y~~~~~~~~~~~~l~~~~~~~~~~~ 159 (217)
T 1gri_A 81 GAFLIRESESAPGDFSLS-VKFGNDVQHFKVLRDGAGKYFLWVVKFNSLNELVDYHRSTSVSRNQQIFLRDIEQVPQQPT 159 (217)
T ss_dssp TCEEEEECSSSTTCEEEE-EEETTEEEEEEEEECSSSCEESSSCEESSHHHHHHHHHHSCSSSSTTCCCCBCCCCCCCCC
T ss_pred CeEEEEECCCCCCCcEEE-EECCCceeEEEEEECCCCcEEEeeEecCCHHHHHHHhhcCCCccccceeccccccccCCce
The seemingly incorrect alignment is explained by the fact that HHsearch / HHblits aligns profile HMMs, not sequences directly. In this example, the query and template multiple sequence alignments (MSAs) from which the profile HMMs were built are not identical, as one can see for instance from the differing Consensus annotation strings and different predicted secondary structure strings. The underying MSAs probably contain the exact same alignment ambiguity in this region, with some member sequences being shifted by one positions between positions 92 and 99 relative to the master sequence of the MSA.
Let’s do a search with the human PIP49/FAM69B protein, for which we
generated an MSA in query.a3m with two iterations of HHblits in
Generating a multiple sequence alignment using HHblits:
Query sp|Q5VUD6|FA69B_HUMAN Protein FAM69B OS=Homo sapiens GN=FAM69B PE=2 SV=3
Match_columns 431
No_of_seqs 149 out of 272
Neff 5.2
Searched_HMMs 13730
Date Wed Jan 4 17:44:24 2012
Command hhsearch -i query.a3m -d /cluster/user/soeding/databases/scop -cpu 18
No Hit Prob E-value P-value Score SS Cols Query HMM Template HMM
1 d1qpca_ d.144.1.7 (A:) Lymphoc 99.7 4.5E-17 3.2E-21 154.3 10.2 99 203-320 56-157 (272)
2 d1jpaa_ d.144.1.7 (A:) ephb2 r 99.7 4.3E-17 3.1E-21 156.8 8.8 99 203-321 75-177 (299)
3 d1uwha_ d.144.1.7 (A:) B-Raf k 99.7 5.1E-17 3.7E-21 154.8 7.7 100 203-322 52-154 (276)
4 d1opja_ d.144.1.7 (A:) Abelson 99.7 6.2E-17 4.5E-21 154.8 8.3 100 203-321 61-164 (287)
5 d1mp8a_ d.144.1.7 (A:) Focal a 99.6 9.9E-17 7.2E-21 151.3 8.6 100 203-322 56-158 (273)
6 d1sm2a_ d.144.1.7 (A:) Tyrosin 99.6 1.2E-16 8.8E-21 150.3 8.8 99 203-321 48-150 (263)
7 d1u59a_ d.144.1.7 (A:) Tyrosin 99.6 2.4E-16 1.7E-20 150.9 9.5 99 203-321 57-158 (285)
8 d1xbba_ d.144.1.7 (A:) Tyrosin 99.6 2.2E-16 1.6E-20 150.2 8.6 97 203-320 56-155 (277)
9 d1vjya_ d.144.1.7 (A:) Type I 99.6 2.6E-16 1.9E-20 151.3 8.8 98 204-320 46-156 (303)
10 d1mqba_ d.144.1.7 (A:) epha2 r 99.6 4.4E-16 3.2E-20 148.0 8.7 193 203-422 57-272 (283)
...
64 d1j7la_ d.144.1.6 (A:) Type II 97.3 0.00014 1E-08 65.0 6.3 33 292-324 184-216 (263)
65 d1nd4a_ d.144.1.6 (A:) Aminogl 96.7 0.0012 8.5E-08 58.5 6.6 31 292-322 176-206 (255)
66 d1nw1a_ d.144.1.8 (A:) Choline 96.6 0.0011 7.8E-08 63.9 5.8 37 203-239 92-128 (395)
67 d2pula1 d.144.1.6 (A:5-396) Me 95.6 0.0071 5.2E-07 58.3 6.4 32 290-322 222-253 (392)
68 d1a4pa_ a.39.1.2 (A:) Calcycli 91.7 0.12 8.9E-06 40.0 5.4 62 140-202 18-80 (92)
69 d1ksoa_ a.39.1.2 (A:) Calcycli 91.2 0.17 1.2E-05 39.5 5.8 56 147-203 28-83 (93)
70 d1e8aa_ a.39.1.2 (A:) Calcycli 90.5 0.23 1.7E-05 38.3 6.0 56 147-203 27-82 (87)
...
175 d1qxpa2 a.39.1.8 (A:515-702) C 23.7 29 0.0021 28.8 3.8 49 137-197 69-118 (188)
176 d1tuza_ a.39.1.7 (A:) Diacylgl 23.5 55 0.004 25.3 5.3 55 143-201 44-106 (118)
177 d1ggwa_ a.39.1.5 (A:) Cdc4p {F 23.1 26 0.0019 27.0 3.2 66 129-197 35-101 (140)
178 d1topa_ a.39.1.5 (A:) Troponin 22.8 72 0.0052 24.5 6.0 58 140-199 65-123 (162)
179 d1otfa_ d.80.1.1 (A:) 4-oxaloc 22.5 66 0.0048 21.5 5.0 40 267-306 12-53 (59)
180 d1oqpa_ a.39.1.5 (A:) Caltract 22.2 32 0.0023 24.0 3.2 32 165-197 3-34 (77)
181 d1df0a1 a.39.1.8 (A:515-700) C 21.7 43 0.0032 27.0 4.5 51 137-199 67-118 (186)
182 d1zfsa1 a.39.1.2 (A:1-93) Calc 21.1 41 0.003 24.6 3.8 30 170-199 8-38 (93)
183 d1snla_ a.39.1.7 (A:) Nucleobi 20.9 23 0.0016 26.2 2.2 24 174-197 18-41 (99)
The summary hit list that is written to the screen shows the best hits from the database, ordered by the probability of being a true positive (column 4: ’Prob’). The meaning of the columns is the following:
No: the index of the datbase match.
Hit: the first 30 characters of the name line.
Prob: the Probability of template to be a true positive.
For the probability of being a true positive, the secondary structure
score in column SS is taken into account, together with the raw score
in column Score. True positives are defined to be either globally
homologous or they are at least homologous in parts, and thereby locally
similar in structure. More precisely, the latter criterion demands that
the MAXSUB score between query and hit is at least 0.1. In almost all
cases the structural similarity will we be due to a global OR LOCAL
homology between query and template.
E-value: The E-value gives the average number of false
positives (’wrong hits’) with a score better than the one for the
template when scanning the database. It is a measure of reliability:
E-values near to 0 signify a very reliable hit, an E-value of 10 means
about 10 wrong hits are expected to be found in the database with a
score at least this good. Note that E-value and P-value are calculated
without taking the secondary structure into account!
P-value: The P-value is the E-value divided by the number
of sequences in the database. It is the probability that in a pairwise
comparison a wrong hit will score at least this good.
Score: the raw score is computed by the Viterbi HMM-HMM
alignment excluding the secondary structure score. It is the sum of
similarities of the aligned profile columns minus the position-specific
gap penalties in bits. The column similarity score is the
log-sum-of-odds score (base 2) as described in the original HHsearch
paper (Soding, Bioinformatics 2005). The gap penalties are the log2 of
the state transition probabilities, e.g. from match state to insert or
delete to match state.
SS: the secondary structure score. This score tells you
how well the PSIPRED-predicted (3-state) or actual DSSP-determined
(8-state) secondary structure sequences agree with each other. PSIPRED
confidence values are used in the scoring, low confidences getting less
statistical weight.
Cols: the number of aligned Match columns in the HMM-HMM
alignment.
Query HMM: the range of aligned match states from the
query HMM.
Template HMM: the range of aligned match states from the
database/template HMM and, in parenthesis, the number of match states in
the database HMM.
The output file d1bpya1.hhr contains the same hit list plus the pairwise HMM alignments. One example is give here:
No 68
>d1a4pa_ a.39.1.2 (A:) Calcyclin (S100) {Human (Homo sapiens), P11 s100a10, calpactin [TaxId: 9606]}
Probab=91.65 E-value=0.12 Score=40.00 Aligned_cols=62 Identities=16% Similarity=0.149 Sum_probs=42.0
Q ss_pred ccCCCCCCcHHHHHHHHHHHHHhhcccCccHHHHHHHHHhhhhccCCCCcCHHHHHHH-HHHHH
Q sp|Q5VUD6|FA69 140 FDKPTRGTSIKEFREMTLSFLKANLGDLPSLPALVGQVLLMADFNKDNRVSLAEAKSV-WALLQ 202 (431)
Q Consensus 140 ~d~p~~g~s~~eF~emv~~~i~~~lg~~~~l~~L~~~~~~~~d~nk~g~vs~~e~~sl-waLlq 202 (431)
||+..-..|.+||.+++.......++.+.+ ...+..++..+|.|+||+|++.|...+ ..|..
T Consensus 18 yd~ddG~is~~El~~~l~~~~~~~~~~~~~-~~~v~~~~~~~D~n~DG~I~F~EF~~li~~l~~ 80 (92)
T d1a4pa_ 18 FAGDKGYLTKEDLRVLMEKEFPGFLENQKD-PLAVDKIMKDLDQCRDGKVGFQSFFSLIAGLTI 80 (92)
T ss_dssp HHGGGCSBCHHHHHHHHHHHCHHHHHHSCC-TTHHHHHHHHHCTTSSSCBCHHHHHHHHHHHHH
T ss_pred HcCCCCEEcHHHHHHHHHHhccccccccCC-HHHHHHHHHHHhCCCCCCCcHHHHHHHHHHHHH
Confidence 444433449999999998876655554332 234566677899999999999997544 44443
This alignment shows an EF hand embedded in a kinase domain in
PIP49/FAM69B. The first line, which begins with with a >, contains
the name and description line of the template/database HMM. (We use
"template HMM" and "matched database HMM" synonymously.) The next line
summarizes the main statistics for the alignment: The probability for
the query and template HMMs to be homologous (Probab), the E-value,
the raw Score, and the number of aligned columns are repeated from the
summary hit list. The Identities give the percentage of aligned
residue pairs of the query and the template master sequences that are
identical. The Similarity is the arithmetic mean of the substitution
scores between the aligned residue pairs from the query and template
master sequences. The substitution matrix is the same as the one used to
calculate the pseudocounts for the database HMMs, by default the Gonnet
matrix. (The matrix can be changed with the -Blosom<XX> option.)
The Sum_probs value is the sum over the posterior probabilities of all
aligned pairs of match states. These probabilities are calculated by the
Forward-Backward algorithm. (They are used by the maximum accuracy
algorithm which computes the final alignments.) When the template HMM
has secondary structure annotation from DSSP, the sum_probs value runs
only over aligned pairs for which the template has a valid DSSP state,
not a - sign. A - would indicate that the structural coordinates of
that residue are missing in the template. For homology modelling, this
special treatment of templates with known structure makes sum_probs a
useful feature to use for ranking templates.
The pairwise alignment consists of one or more blocks with the following lines:
Q ss_dssp: the query secondary structure as determined by DSSP (when available)
Q ss_pred: the query secondary structure as predicted by PSIPRED (when available)
Q <Q_name>: the query master sequence
Q Consensus: the query alignment consensus sequence
The predicted secondary structure states are shown in capital letters if
the PSIPRED confidence value is between 0.7 and 1.0, for lower
confidence values they are given in lower-case letters. With the option
-ssconf, ss_conf lines can be added to
the alignments which report the PSIPRED confidence values by numbers
between 0 and 9 (as in versions up to 1.5).
The consensus sequence uses capital letters for well conserved columns
and lower case for partially conserved columns. Unconserved columns are
marked by a tilde ~. Roughly speaking, amino acids that occur with
(>+ 60%) probability (before adding pseudocounts) are written as
capital letters and amino acids that have (>= 40%) probability are
written as lower case letters, where gaps are included in the fraction
counts. More precisely, when the gap-corrected amino acid fraction
p_i(a) * N_eff(i)/(N_eff + 1)
is above 0.6 (0.4) an upper (lower) case letter is used for amino acid a. Here, p_i(a) is
the emission probability for a in column i, N_eff is the
effective number of sequences in the entire multiple alignment (between
1 and 20) and N_eff(i) is the effective number of sequences
in the subalignment consisting of those sequences that do not have a gap
in column i. These percentages increase approximately inversely
proportionally with the fraction of gaps in the column, hence a column
with only cysteines and 50% gaps gets a lower case letter.
The line in the middle shows the column score between the query and template amino acid distributions. It gives a valuable indication for the alignment quality.
= : column score below -1.5
- : column score between -1.5 and -0.5
. : column score between -0.5 and +0.5
+ : column score between +0.5 and +1.5
| : column score above +1.5
A unit of column score corresponds approximately to 0.6 bits. From the
column score line the excellent alignment around the conserved
D.n.DG.i...E motif in the turn between two helices is
evident. The alignment around the gap by contrast scores only a bit
better than zero per residue and is therefore not very reliable.
After the template block, which consists of the following lines,
T Consensus: the template alignment consensus sequence
T <T_name>: the template domain sequence
T ss_dssp: the template secondary structure as determined by DSSP (when available)
T ss_pred: the template secondary structure as predicted by PSIPRED (when available)
The last line in the block (Confidence) reports the
reliability of the pairwise query-template alignment. The confidence
values are obtained from the posterior probabilities calculated in the
Forward-Backward algorithm. A value of 8 indicates a probability that
this pair of HMM columns is correctly aligned between 0.8 and 0.9. The
Confidence line is only displayed when the -realign option
is active.
Multiple alignments can be read in A2M, A3M, or aligned FASTA format.
(Check the -M option for using an input format different from the
default A3M). You can transform MSAs from Clustal or Stockholm format to
A3M or aligned FASTA with the reformat.pl utility supplied in this
package.
To reformat from Clustal format to A3M:
reformat.pl test.aln test.a3m
or explicitly, if the formats can not be recognized from the extensions:
reformat.pl clu a3m test.clustal test.a3m
To reformat from Stockholm to aligned FASTA:
reformat.pl test.sto test.fas
>d1a1x__ b.63.1.1 (-) p13-MTCP1 {Human (Homo sapiens)}
PPDHLWVHQEGIYRDEYQRTWVAVVEE--E--T--SF---------LR----------ARVQQIQVPLG-------DAARPSHLLTS-----QL
>gi|6678257|ref|NP_033363.1|:(7-103) T-cell lymphoma breakpoint 1 [Mus musculus]
HPNRLWIWEKHVYLDEFRRSWLPVVIK--S--N--EK---------FQ----------VILRQEDVTLG-------EAMSPSQLVPY-----EL
>gi|7305557|ref|NP_038800.1|:(8-103) T-cell leukemia/lymphoma 1B, 3 [Mus musculus]
PPRFLVCTRDDIYEDENGRQWVVAKVE--T--S--RSpygsrietcIT----------VHLQHMTTIPQ-------EPTPQQPINNN-----SL
>gi|11415028|ref|NP_068801.1|:(2-106) T-cell lymphoma-1; T-cell lymphoma-1A [Homo sapiens]
HPDRLWAWEKFVYLDEKQHAWLPLTIEikD--R--LQ---------LR----------VLLRREDVVLG-------RPMTPTQIGPS-----LL
>gi|7305561|ref|NP_038804.1|:(7-103) T-cell leukemia/lymphoma 1B, 5 [Mus musculus]
----------GIYEDEHHRVWIAVNVE--T--S--HS---------SHgnrietcvt-VHLQHMTTLPQ-------EPTPQQPINNN-----SL
>gi|7305553|ref|NP_038801.1|:(5-103) T-cell leukemia/lymphoma 1B, 1 [Mus musculus]
LPVYLVSVRLGIYEDEHHRVWIVANVE--TshS--SH---------GN----------RRRTHVTVHLW-------KLIPQQVIPFNplnydFL
>gi|27668591|ref|XP_234504.1|:(7-103) similar to Chain A, Crystal Structure Of Murine Tcl1
-PDRLWLWEKHVYLDEFRRSWLPIVIK--S--N--GK---------FQ----------VIMRQKDVILG-------DSMTPSQLVPY-----EL
>gi|27668589|ref|XP_234503.1|:(9-91) similar to T-cell leukemia/lymphoma 1B, 5;
-PHILTLRTHGIYEDEHHRLWVVLDLQ--A--ShlSF---------SN----------RLLIYLTVYLQqgvafplESTPPSPMNLN-----GL
>gi|7305559|ref|NP_038802.1|:(8-102) T-cell leukemia/lymphoma 1B, 4 [Mus musculus]
PPCFLVCTRDDIYEDEHGRQWVAAKVE--T--S--SH---------SPycskietcvtVHLWQMTTLFQ-------EPSPDSLKTFN-----FL
>gi|7305555|ref|NP_038803.1|:(9-102) T-cell leukemia/lymphoma 1B, 2 [Mus musculus]
---------PGFYEDEHHRLWMVAKLE--T--C--SH---------SPycnkietcvtVHLWQMTRYPQ-------EPAPYNPMNYN-----FL
The sequence name and its description must be contained in a single name
line beginning with the > symbol and followed directly by the
sequence name. The residue data is contained in one or more lines of
arbitrary length following the name line. No empty lines should be used.
In aligned FASTA the gaps are written with - and the n'th letter of
each sequence (except newlines) is understood to build the n'th column
of the multiple alignment.
>d1a1x__ b.63.1.1 (-) p13-MTCP1 {Human (Homo sapiens)}
PPDHLWVHQEGIYRDEYQRTWVAVVEE..E..T..SF.........LR..........ARVQQIQVPLG.......DAARPSHLLTS.....QL
>gi|6678257|ref|NP_033363.1|:(7-103) T-cell lymphoma breakpoint 1 [Mus musculus]
HPNRLWIWEKHVYLDEFRRSWLPVVIK..S..N..EK.........FQ..........VILRQEDVTLG.......EAMSPSQLVPY.....EL
>gi|7305557|ref|NP_038800.1|:(8-103) T-cell leukemia/lymphoma 1B, 3 [Mus musculus]
PPRFLVCTRDDIYEDENGRQWVVAKVE..T..S..RSpygsrietcIT..........VHLQHMTTIPQ.......EPTPQQPINNN.....SL
>gi|11415028|ref|NP_068801.1|:(2-106) T-cell lymphoma-1; T-cell lymphoma-1A [Homo sapiens]
HPDRLWAWEKFVYLDEKQHAWLPLTIEikD..R..LQ.........LR..........VLLRREDVVLG.......RPMTPTQIGPS.....LL
>gi|7305561|ref|NP_038804.1|:(7-103) T-cell leukemia/lymphoma 1B, 5 [Mus musculus]
----------GIYEDEHHRVWIAVNVE..T..S..HS.........SHgnrietcvt.VHLQHMTTLPQ.......EPTPQQPINNN.....SL
>gi|7305553|ref|NP_038801.1|:(5-103) T-cell leukemia/lymphoma 1B, 1 [Mus musculus]
LPVYLVSVRLGIYEDEHHRVWIVANVE..TshS..SH.........GN..........RRRTHVTVHLW.......KLIPQQVIPFNplnydFL
>gi|27668591|ref|XP_234504.1|:(7-103) similar to Chain A, Crystal Structure Of Murine Tcl1
-PDRLWLWEKHVYLDEFRRSWLPIVIK..S..N..GK.........FQ..........VIMRQKDVILG.......DSMTPSQLVPY.....EL
>gi|27668589|ref|XP_234503.1|:(9-91) similar to T-cell leukemia/lymphoma 1B, 5;
-PHILTLRTHGIYEDEHHRLWVVLDLQ..A..ShlSF.........SN..........RLLIYLTVYLQqgvafplESTPPSPMNLN.....GL
>gi|7305559|ref|NP_038802.1|:(8-102) T-cell leukemia/lymphoma 1B, 4 [Mus musculus]
PPCFLVCTRDDIYEDEHGRQWVAAKVE..T..S..SH.........SPycskietcvtVHLWQMTTLFQ.......EPSPDSLKTFN.....FL
>gi|7305555|ref|NP_038803.1|:(9-102) T-cell leukemia/lymphoma 1B, 2 [Mus musculus]
---------PGFYEDEHHRLWMVAKLE..T..C..SH.........SPycnkietcvtVHLWQMTRYPQ.......EPAPYNPMNYN.....FL
A2M format is derived from aligned FASTA format. It looks very similar,
but it distinguishes between match/delete columns and insert columns.
This information is important to uniquely specify how an alignment is
transformed into an HMM. The match/delete columns use upper case letters
for residues and the - symbol for deletions (gaps). The insert columns
use lower case letters for the inserted residues. Gaps aligned to
inserted residues are written as . Lines beginning with a hash #
symbol will be treated as commentary lines in HHsearch/HHblits (see
below).
>d1a1x__ b.63.1.1 (-) p13-MTCP1 {Human (Homo sapiens)}
PPDHLWVHQEGIYRDEYQRTWVAVVEEETSFLRARVQQIQVPLGDAARPSHLLTSQL
>gi|6678257|ref|NP_033363.1|:(7-103) T-cell lymphoma breakpoint 1 [Mus musculus]
HPNRLWIWEKHVYLDEFRRSWLPVVIKSNEKFQVILRQEDVTLGEAMSPSQLVPYEL
>gi|7305557|ref|NP_038800.1|:(8-103) T-cell leukemia/lymphoma 1B, 3 [Mus musculus]
PPRFLVCTRDDIYEDENGRQWVVAKVETSRSpygsrietcITVHLQHMTTIPQEPTPQQPINNNSL
>gi|11415028|ref|NP_068801.1|:(2-106) T-cell lymphoma-1; T-cell lymphoma-1A [Homo sapiens]
HPDRLWAWEKFVYLDEKQHAWLPLTIEikDRLQLRVLLRREDVVLGRPMTPTQIGPSLL
>gi|7305561|ref|NP_038804.1|:(7-103) T-cell leukemia/lymphoma 1B, 5 [Mus musculus]
----------GIYEDEHHRVWIAVNVETSHSSHgnrietcvtVHLQHMTTLPQEPTPQQPINNNSL
>gi|7305553|ref|NP_038801.1|:(5-103) T-cell leukemia/lymphoma 1B, 1 [Mus musculus]
LPVYLVSVRLGIYEDEHHRVWIVANVETshSSHGNRRRTHVTVHLWKLIPQQVIPFNplnydFL
>gi|27668591|ref|XP_234504.1|:(7-103) similar to Chain A, Crystal Structure Of Murine Tcl1
-PDRLWLWEKHVYLDEFRRSWLPIVIKSNGKFQVIMRQKDVILGDSMTPSQLVPYEL
>gi|27668589|ref|XP_234503.1|:(9-91) similar to T-cell leukemia/lymphoma 1B, 5;
-PHILTLRTHGIYEDEHHRLWVVLDLQAShlSFSNRLLIYLTVYLQqgvafplESTPPSPMNLNGL
>gi|7305559|ref|NP_038802.1|:(8-102) T-cell leukemia/lymphoma 1B, 4 [Mus musculus]
PPCFLVCTRDDIYEDEHGRQWVAAKVETSSHSPycskietcvtVHLWQMTTLFQEPSPDSLKTFNFL
>gi|7305555|ref|NP_038803.1|:(9-102) T-cell leukemia/lymphoma 1B, 2 [Mus musculus]
---------PGFYEDEHHRLWMVAKLETCSHSPycnkietcvtVHLWQMTRYPQEPAPYNPMNYNFL
The A3M format is a condensed version of A2M format. It is obtained by
omitting all . symbols from A2M format. Hence residues emitted by
Match states of the HMM are in upper case, residues emitted by Insert
states are in lower case and deletions are written -. A3M-formatted
alignments can be reformatted to other formats like FASTA or A2M with
the reformat.pl utility:
reformat.pl test.a3m test.a2m
Lines beginning with a hash # symbol will be treated as commentary
lines in HHsearch/HHblits (see below). Please note that A3M, though very
practical and space-efficient, is not a standard format, and the name
A3M is our personal
invention.
The alignments read in by HHblits, HHsearch or HHmake can also contain secondary structure information. This information can be included in sequences with special names, like in this A3M file:
>ss_dssp
CCSEEEEEETTEEEETTSCEEEEEEEECSSCEEEEEECCCCCCCSCCCHHHHTTCSSCSEEEEETTTEEEETTSC
>aa_dssp
PPDHLWVHQEGIYRDEYQRTWVAVVEEETSFLRARVQQIQVPLGDAARPSHLLTSQLPLMWQLYPEERYMDNNSR
>aa_pred
PPDHLWVHQEGIYRDEYQRTWVAVVEEETSFLRARVQQIQVPLGDAARPSHLLTSQLPLMWQLYPEERYMDNNSR
>ss_pred
CCCEEEEECCCEECCCCCEEEEEEEEECCCCCCEEEEEEECCCCCCCCCCCCCCCCCCCEEEECCCCCEECCCCC
>ss_conf
987689961870104587078999970578640132153103788788777774424614787217702035631
>d1a1x__ b.63.1.1 (-) p13-MTCP1 {Human (Homo sapiens)}
PPDHLWVHQEGIYRDEYQRTWVAVVEEETSFLRARVQQIQVPLGDAARPSHLLTSQLPLMWQLYPEERYMDNNSR
>gi|6678257|ref|NP_033363.1|:(7-103) T-cell lymphoma breakpoint 1 [Mus musculus]
HPNRLWIWEKHVYLDEFRRSWLPVVIKSNEKFQVILRQEDVTLGEAMSPSQLVPYELPLMWQLYPKDRYRSCDSM
>gi|7305557|ref|NP_038800.1|:(8-103) T-cell leukemia/lymphoma 1B, 3 [Mus musculus]
PPRFLVCTRDDIYEDENGRQWVVAKVETSRSpygsrietcITVHLQHMTTIPQEPTPQQPINNNSLPTMWRLESMNTYTGTDGT
>gi|11415028|ref|NP_068801.1|:(2-106) T-cell lymphoma-1; T-cell lymphoma-1A [Homo sapiens]
HPDRLWAWEKFVYLDEKQHAWLPLTIEikDRLQLRVLLRREDVVLGRPMTPTQIGPSLLPIMWQLYPDGRYRSSDSS
The sequence with name >ss_dssp contains the 8-state DSSP-determined
secondary structure. >aa_dssp and >aa_pred contain the same residues
as the query sequence (>d1a1x__ in this case). They are optional and
used merely to check whether the secondary structure states have
correctly been assigned to the alignment. >ss_pred contains the
3-state secondary structure predicted by PSIPRED, and >ss_conf
contains the corresponding confidence values. The query sequence is the
first sequence that does not start with a special name. It is not marked
explicitly.
If you would like to create HMMs from alignments with a specified name which differ from the name of the first sequence, you can do so by adding name lines to your FASTA, A2M, or A3M alignment:
#PF02043 Bac_chlorC: Bacteriochlorophyll C binding protein
>ss_pred
CCCCHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHHCCCCCCCCCCCCCCCCCCCCCCCCCCCCHHHHHHHCC
>ss_conf
9863234658887677777750019999999998878886403445886666775655576678777660667633039
>CSMA_CHLAU/1-79
ATRGWFSESSAQVAQIGDIMFQGHWQWVSNALQATAAAVDNINRNAYPGVSRSGSGEGAFSSSPSNGFRPKRIRSRFNR
>CSMA_CHLPH/2-51
NGGGVFTDILAASGRIFEVMVEGHWATVGYLFDSLGKGVSRINQNAYGNM-----------------------------
...
When creating an HMM from an A3M file with hhmake, the first word of the name line is used as the name and file name of the HMM (PF02043 in this case). The following is an optional description. The descriptions will appear in the hit list and alignment section of the search results. The name lines can be arbitrarily long and there can be any number of name/description lines included, marked by a ’#’ as the first character in the line. Note that name lines are read by HHmake but are not a part of the standard definition of the FASTA or A2M format.
HMMs can be read by HHsearch/HHblits in HHsuite’s own hhm format.
Performance is severely reduced when using HMMER-format, because HMMER
adds more pseudocounts than HHsearch/HHblits that we cannot remove
anymore. We therefore strongly advise against using models from HMMER
with HHsuite. We recommend to get the original MSA files and generate
hhm-formatted models using hhmake, as described in subsection
Generating a multiple sequence alignment using HHblits.
If you absolutely need to use HMMER format, you can convert it to hhm
format with hhmake:
hhmake -i test.hmm -o test.hhm
This works only for a single HMM per file, not for concatenated HMMs.
You may add predicted secondary structure to the hmm file with
addss.pl as for hhm format.
HHsearch/HHblits uses a format HMM that is unchanged since HHsearch version 1.5. This is the example of an HHM model file produced by HHmake:
HHsearch 1.5
NAME d1mvfd_ b.129.1.1 (D:) MazE {Escherichia coli}
FAM b.129.1.1
FILE d1mvfd_
COM hhmake1 -i d1mvfd_.a3m -o test.hhm
DATE Wed May 14 10:41:06 2011
LENG 44 match states, 44 columns in multiple alignment
FILT 32 out of 35 sequences passed filter (-id 90 -cov 0 -qid 0 -qsc -20.00 -diff 100)
NEFF 4.0
SEQ
>ss_dssp
CBCEEETTEEEEECCHHHHHHTTCCTTCBEEEEEETTEEEEEEC
>ss_pred
CCCCCCCCCCCCCCHHHHHHHHCCCCCCEEEEEEECCEEEEEEC
>ss_conf
93233467666600578899808998986889874993798739
>Consensus
sxIxKWGNSxAvRlPaxlxxxlxlxxgdxixxxxxxxxivlxPv
>d1mvfd_ b.129.1.1 (D:) MazE {Escherichia coli}
SSVKRWGNSPAVRIPATLMQALNLNIDDEVKIDLVDGKLIIEPV
>gi|10176344|dbj|BAB07439.1|:(1-43) suppressor of ppGpp-regulated growth inhibitor [Bacillus halodurans]
TTIQKWGNSLAVRIPNHYAKHINVTQGSEIELSLgSDQTIILKP-
>gi|50120611|ref|YP_049778.1|:(3-43) suppressor of growth inhibitory protein ChpA [Erwinia carotovora]
-TVKKWGNSPAIRLSSSVMQAFDMTFNDSFDMEIRETEIALIP-
>gi|44064461|gb|EAG93225.1|:(2-42) unknown [environmental sequence]
-SVVKWGSYLAVRLPAELVLELGLKEGDEIDLVKDDGPVRVR--
>gi|31442758|gb|AAP55635.1|:(1-44) PemI-like protein [Pediococcus acidilactici]
TRLAKWGNSKAARIPSQIIKQLKLDDNQDMTITIENGSIVLTPI
>gi|44419085|gb|EAJ13619.1|:(3-43) unknown [environmental sequence]
SAIQKWGNSAAVRLPAVLLEQIDASVGSSLNADVRPDGVLLSP-
>gi|24376549|gb|AAN57947.1|:(3-44) putative cell growth regulatory protein [Streptococcus mutans UA159]
SAINKWGNSSAIRLPKQLVQELQLQTNDVLDYKVSGNKIILEKV
>gi|11344928|gb|AAG34554.1|:(1-44) MazE [Photobacterium profundum]
TQIRKIGNSLGSIIPATFIRQLELAEGAEIDVKTVDGKIVIEPI
>gi|45681193|ref|ZP_00192636.1|:(2-44) COG2336: Growth regulator [Mesorhizobium sp. BNC1]
-TIRKIGNSEGVILPKELLDRHNLKTGDALAIVEEGSDLVLKPV
#
NULL 3706 5728 4211 4064 4839 3729 4763 4308 4069 3323 5509 4640 4464 4937 4285 4423 3815 3783 6325 4665
HMM A C D E F G H I K L M N P Q R S T V W Y
M->M M->I M->D I->M I->I D->M D->D Neff NeffI NeffD
0 * * 0 * 0 * * * *
S 1 * * * * * * * * * * * * * * * 1012 988 * * * 1
0 * * * * * * 2817 0 0
S 2 2307 * * * * * * * * * * * * 3178 3009 2179 1546 * * * 2
0 * * * * * * 3447 0 0
V 3 * * * * * * * 917 * 3009 * * * * * * * 1530 * * 3
0 * * * * * * 3447 0 0
.
.
.
V 44 * * * * * * * 1309 * * * * * * * * * 745 * * 44
0 * * 0 * * * 2533 0 0
//
The first line (HHsearch 1.5) gives the format version, which
corresponds to the HHsearch version for which this format was first
introduced. Newer versions of HHsearch/HHblits may use previous format
versions. The NAME line gives the name of the HMM and an optional
description. The first 30 characters of this field are used in the
summary hit list of the search results in hhr format, the full name line
is given above the query-template alignments of the search results. The
FAM line contains the family if the sequence is from SCOP of PFAM
(used for calibration). COM is the command that was used to generate
the file. NEFF is the diversity of the alignment, calculated as exp of the negative entropy averaged over all columns of the
alignment.
The SEQ section contains a number of aligned, representative (pseudo)
sequences in A3M format and is terminated with a line containing only a
#. The first sequence represents the DSSP secondary structure (if
available, i.e., if contained in the A3M or FASTA alignment from which
the HMM model was built), the second and third sequences contain the
predicted secondary structure and the corresponding confidence values in
the range 0–9 (if available). The fourth sequence is the consensus
annotation sequence that is shown in the pairwise query-template
alignments in the hhsearch output. The first real sequence after the
pseudo sequences is the seed or master sequence from which the
alignment was built (>d1mvfd_, in our example). If the alignment does
not represent a single master sequence but an entire family, as in the
case of PFAM alignments for example, the first real sequence may be a
consensus sequence calculated for the entire alignment. This master
sequence is shown in the pairwise query-template alignments in the
hhsearch output.
The next line specifies the null model frequencies, which are extracted from the selected substitution matrix used to add pseudocounts. Each of the positive integers is equal to 1000 times the negative logarithm of the amino acid frequency (which is between 0 and 1):
-1000 * log_2(frequency)
After the two annotation lines that specify the order of columns for the emission and transition probabilities that follow, there is a line which is not currently read by HHsearch and that lists the transition frequencies from the begin state to the first Match state, Insert state and Delete state.
The last block contains two lines for each column of the HMM. The first
line starts with the amino acid in the master sequence at that column in
the HMM and the column number. Following are 20 positive integers
representing the match state amino acid emission frequencies (see
equation above). Asterisks * stand for a
frequency of 0 (which would otherwise be represented by 99999). Please
note that, unlike in HMMER format, the emission frequencies do not
contain pseudo-counts in the HHsearch model format. The second line
contains the seven transition frequencies coded as above. The three local diversities,
Neff_M, Neff_I, and Neff_D are given in units of 0.001 (see next
paragraph). The end of the model is indicated by a line containing only
//.
This is just a brief summary of command line parameters for the various
binaries and perl scripts as they are displayed by the programs when
calling them without command line parameters. On the help pages of the
HHpred/HHblits web server you can find
more detailed explanations about some of the input parameters
(Parameters section) and about how to interpret the output
(Results section). The FAQ section contains valuable practical
hints on topics such as how to validate marginally significant database
matches or how to avoid high-scoring false
positives.
HHblits is a sensitive, general-purpose, iterative sequence search tool that represents both query and database sequences by HMMs. You can search HHsuite databases starting with a single query sequence, a multiple sequence alignment (MSA), or an HMM. HHblits prints out a ranked list of database HMMs/MSAs and can also generate an MSA by merging the significant database HMMs/MSAs onto the query MSA.
The binary hhblits_omp supports the parallelization over several
queries with OpenMP. In this case the input needs to be the basename of
an ffindex with MSA’s. The outputs are treated as the basenames for
output ffindices.
Assume the ffindex database
/home/user/databases/scop_a3m.ff{data,index}, the corresponding
basename is /home/user/databases/scop_a3m
Usage: hhblits -i query [options]
-i <file> input/query: single sequence or multiple sequence alignment (MSA)
in a3m, a2m, or FASTA format, or HMM in hhm format
<file> may be 'stdin' or 'stdout' throughout.
Options:
-d <name> database name (e.g. uniclust30_2018_08)
Multiple databases may be specified with '-d <db1> -d <db2> ...'
-n [1,8] number of iterations (default=2)
-e [0,1] E-value cutoff for inclusion in result alignment (def=0.001)
Input alignment format:
-M a2m use A2M/A3M (default): upper case = Match; lower case = Insert;
' -' = Delete; '.' = gaps aligned to inserts (may be omitted)
-M first use FASTA: columns with residue in 1st sequence are match states
-M [0,100] use FASTA: columns with fewer than X% gaps are match states
-tags/-notags do NOT / do neutralize His-, C-myc-, FLAG-tags, and trypsin
recognition sequence to background distribution (def=-notags)
Output options:
-o <file> write results in standard format to file (default=<infile.hhr>)
-oa3m <file> write result MSA with significant matches in a3m format
-opsi <file> write result MSA of significant matches in PSI-BLAST format
-ohhm <file> write HHM file for result MSA of significant matches
-oalis <name> write MSAs in A3M format after each iteration
-add_cons generate consensus sequence as master sequence of query MSA (default=don't)
-hide_cons don't show consensus sequence in alignments (default=show)
-hide_pred don't show predicted 2ndary structure in alignments (default=show)
-hide_dssp don't show DSSP 2ndary structure in alignments (default=show)
-show_ssconf show confidences for predicted 2ndary structure in alignments
-Ofas <file> write pairwise alignments in FASTA xor A2M (-Oa2m) xor A3M (-Oa3m) format
-seq <int> max. number of query/template sequences displayed (default=1)
-aliw <int> number of columns per line in alignment list (default=80)
-p [0,100] minimum probability in summary and alignment list (default=20)
-E [0,inf[ maximum E-value in summary and alignment list (default=1E+06)
-Z <int> maximum number of lines in summary hit list (default=500)
-z <int> minimum number of lines in summary hit list (default=10)
-B <int> maximum number of alignments in alignment list (default=500)
-b <int> minimum number of alignments in alignment list (default=10)
Prefilter options
-noprefilt disable all filter steps
-noaddfilter disable all filter steps (except for fast prefiltering)
-maxfilt max number of hits allowed to pass 2nd prefilter (default=20000)
-min_prefilter_hits min number of hits to pass prefilter (default=100)
-prepre_smax_thresh min score threshold of ungapped prefilter (default=10)
-pre_evalue_thresh max E-value threshold of Smith-Waterman prefilter score (default=1000.0)
-pre_bitfactor prefilter scores are in units of 1 bit / pre_bitfactor (default=4)
-pre_gap_open gap open penalty in prefilter Smith-Waterman alignment (default=20)
-pre_gap_extend gap extend penalty in prefilter Smith-Waterman alignment (default=4)
-pre_score_offset offset on sequence profile scores in prefilter S-W alignment (default=50)
Filter options applied to query MSA, database MSAs, and result MSA
-all show all sequences in result MSA; do not filter result MSA
-id [0,100] maximum pairwise sequence identity (def=90)
-diff [0,inf[ filter MSAs by selecting most diverse set of sequences, keeping
at least this many seqs in each MSA block of length 50
Zero and non-numerical values turn off the filtering. (def=1000)
-cov [0,100] minimum coverage with master sequence (%) (def=0)
-qid [0,100] minimum sequence identity with master sequence (%) (def=0)
-qsc [0,100] minimum score per column with master sequence (default=-20.0)
-neff [1,inf] target diversity of multiple sequence alignment (default=off)
-mark do not filter out sequences marked by ">@"in their name line
HMM-HMM alignment options:
-norealign do NOT realign displayed hits with MAC algorithm (def=realign)
-realign_old_hits realign hits from previous iterations
-mact [0,1[ posterior prob threshold for MAC realignment controlling greedi-
ness at alignment ends: 0:global >0.1:local (default=0.35)
-glob/-loc use global/local alignment mode for searching/ranking (def=local)
-realign realign displayed hits with max. accuracy (MAC) algorithm
-realign_max <int> realign max. <int> hits (default=500)
-ovlp <int> banded alignment: forbid <ovlp> largest diagonals |i-j| of DP matrix (def=0)
-alt <int> show up to this many alternative alignments with raw score > smin(def=4)
-smin <float> minimum raw score for alternative alignments (def=20.0)
-shift [-1,1] profile-profile score offset (def=-0.03)
-corr [0,1] weight of term for pair correlations (def=0.10)
-sc <int> amino acid score (tja: template HMM at column j) (def=1)
0 = log2 Sum(tja*qia/pa) (pa: aa background frequencies)
1 = log2 Sum(tja*qia/pqa) (pqa = 1/2*(pa+ta) )
2 = log2 Sum(tja*qia/ta) (ta: av. aa freqs in template)
3 = log2 Sum(tja*qia/qa) (qa: av. aa freqs in query)
5 local amino acid composition correction
-ssm {0,..,4} 0: no ss scoring
1,2: ss scoring after or during alignment [default=2]
3,4: ss scoring after or during alignment, predicted vs. predicted
-ssw [0,1] weight of ss score (def=0.11)
-ssa [0,1] ss confusion matrix = (1-ssa)*I + ssa*psipred-confusion-matrix [def=1.00)
-wg use global sequence weighting for realignment!
Gap cost options:
-gapb [0,inf[ Transition pseudocount admixture (def=1.00)
-gapd [0,inf[ Transition pseudocount admixture for open gap (default=0.15)
-gape [0,1.5] Transition pseudocount admixture for extend gap (def=1.00)
-gapf ]0,inf] factor to increase/reduce gap open penalty for deletes (def=0.60)
-gapg ]0,inf] factor to increase/reduce gap open penalty for inserts (def=0.60)
-gaph ]0,inf] factor to increase/reduce gap extend penalty for deletes(def=0.60)
-gapi ]0,inf] factor to increase/reduce gap extend penalty for inserts(def=0.60)
-egq [0,inf[ penalty (bits) for end gaps aligned to query residues (def=0.00)
-egt [0,inf[ penalty (bits) for end gaps aligned to template residues (def=0.00)
Pseudocount (pc) options:
Context specific hhm pseudocounts:
-pc_hhm_contxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=2)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
3: CSBlast admixture: tau = a(1+b)/(Neff[i]+b)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_hhm_contxt_a [0,1] overall pseudocount admixture (def=0.9)
-pc_hhm_contxt_b [1,inf[ Neff threshold value for mode 2 (def=4.0)
-pc_hhm_contxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context independent hhm pseudocounts (used for templates; used for query if contxt file is not available):
-pc_hhm_nocontxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=2)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_hhm_nocontxt_a [0,1] overall pseudocount admixture (def=1.0)
-pc_hhm_nocontxt_b [1,inf[ Neff threshold value for mode 2 (def=1.5)
-pc_hhm_nocontxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context specific prefilter pseudocounts:
-pc_prefilter_contxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=3)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
3: CSBlast admixture: tau = a(1+b)/(Neff[i]+b)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_prefilter_contxt_a [0,1] overall pseudocount admixture (def=0.8)
-pc_prefilter_contxt_b [1,inf[ Neff threshold value for mode 2 (def=2.0)
-pc_prefilter_contxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context independent prefilter pseudocounts (used if context file is not available):
-pc_prefilter_nocontxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=2)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_prefilter_nocontxt_a [0,1] overall pseudocount admixture (def=1.0)
-pc_prefilter_nocontxt_b [1,inf[ Neff threshold value for mode 2 (def=1.5)
-pc_prefilter_nocontxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context-specific pseudo-counts:
-nocontxt use substitution-matrix instead of context-specific pseudocounts
-contxt <file> context file for computing context-specific pseudocounts (default=${HHLIB}/data/context_data.crf)
-csw [0,inf] weight of central position in cs pseudocount mode (def=1.6)
-csb [0,1] weight decay parameter for positions in cs pc mode (def=0.9)
Other options:
-v <int> verbose mode: 0:no screen output 1:only warings 2: verbose (def=2)
-neffmax ]1,20] skip further search iterations when diversity Neff of query MSA
becomes larger than neffmax (default=10.0)
-cpu <int> number of CPUs to use (for shared memory SMPs) (default=2)
-scores <file> write scores for all pairwise comparisons to file
-atab <file> write all alignments in tabular layout to file
-maxres <int> max number of HMM columns (def=20001)
-maxmem [1,inf[ limit memory for realignment (in GB) (def=3.0)
Example: hhblits -i query.fas -oa3m query.a3m -n 1
Usage: hhsearch -i query -d database [options]
-i <file> input/query multiple sequence alignment (a2m, a3m, FASTA) or HMM
<file> may be 'stdin' or 'stdout' throughout.
Options:
-d <name> database name (e.g. pdb70)
Multiple databases may be specified with '-d <db1> -d <db2> ...'
-e [0,1] E-value cutoff for inclusion in result alignment (def=0.001)
Input alignment format:
-M a2m use A2M/A3M (default): upper case = Match; lower case = Insert;
'-' = Delete; '.' = gaps aligned to inserts (may be omitted)
-M first use FASTA: columns with residue in 1st sequence are match states
-M [0,100] use FASTA: columns with fewer than X% gaps are match states
-tags/-notags do NOT / do neutralize His-, C-myc-, FLAG-tags, and trypsin
recognition sequence to background distribution (def=-notags)
Output options:
-o <file> write results in standard format to file (default=<infile.hhr>)
-oa3m <file> write result MSA with significant matches in a3m format
-opsi <file> write result MSA of significant matches in PSI-BLAST format
-ohhm <file> write HHM file for result MSA of significant matches
-add_cons generate consensus sequence as master sequence of query MSA (default=don't)
-hide_cons don't show consensus sequence in alignments (default=show)
-hide_pred don't show predicted 2ndary structure in alignments (default=show)
-hide_dssp don't show DSSP 2ndary structure in alignments (default=show)
-show_ssconf show confidences for predicted 2ndary structure in alignments
-Ofas <file> write pairwise alignments in FASTA xor A2M (-Oa2m) xor A3M (-Oa3m) format
-seq <int> max. number of query/template sequences displayed (default=1)
-aliw <int> number of columns per line in alignment list (default=80)
-p [0,100] minimum probability in summary and alignment list (default=20)
-E [0,inf[ maximum E-value in summary and alignment list (default=1E+06)
-Z <int> maximum number of lines in summary hit list (default=500)
-z <int> minimum number of lines in summary hit list (default=10)
-B <int> maximum number of alignments in alignment list (default=500)
-b <int> minimum number of alignments in alignment list (default=10)
Filter options applied to query MSA, database MSAs, and result MSA
-all show all sequences in result MSA; do not filter result MSA
-id [0,100] maximum pairwise sequence identity (def=90)
-diff [0,inf[ filter MSAs by selecting most diverse set of sequences, keeping
at least this many seqs in each MSA block of length 50
Zero and non-numerical values turn off the filtering. (def=100)
-cov [0,100] minimum coverage with master sequence (%) (def=0)
-qid [0,100] minimum sequence identity with master sequence (%) (def=0)
-qsc [0,100] minimum score per column with master sequence (default=-20.0)
-neff [1,inf] target diversity of multiple sequence alignment (default=off)
-mark do not filter out sequences marked by ">@"in their name line
HMM-HMM alignment options:
-norealign do NOT realign displayed hits with MAC algorithm (def=realign)
-ovlp <int> banded alignment: forbid <ovlp> largest diagonals |i-j| of DP matrix (def=0)
-mact [0,1[ posterior prob threshold for MAC realignment controlling greedi-
ness at alignment ends: 0:global >0.1:local (default=0.35)
-glob/-loc use global/local alignment mode for searching/ranking (def=local)
-realign realign displayed hits with max. accuracy (MAC) algorithm
-excl <range> exclude query positions from the alignment, e.g. '1-33,97-168'
-realign_max <int> realign max. <int> hits (default=500)
-alt <int> show up to this many alternative alignments with raw score > smin(def=4)
-smin <float> minimum raw score for alternative alignments (def=20.0)
-shift [-1,1] profile-profile score offset (def=-0.03)
-corr [0,1] weight of term for pair correlations (def=0.10)
-sc <int> amino acid score (tja: template HMM at column j) (def=1)
0 = log2 Sum(tja*qia/pa) (pa: aa background frequencies)
1 = log2 Sum(tja*qia/pqa) (pqa = 1/2*(pa+ta) )
2 = log2 Sum(tja*qia/ta) (ta: av. aa freqs in template)
3 = log2 Sum(tja*qia/qa) (qa: av. aa freqs in query)
5 local amino acid composition correction
-ssm {0,..,4} 0: no ss scoring
1,2: ss scoring after or during alignment [default=2]
3,4: ss scoring after or during alignment, predicted vs. predicted
-ssw [0,1] weight of ss score (def=0.11)
-ssa [0,1] SS substitution matrix = (1-ssa)*I + ssa*full-SS-substition-matrix [def=1.00)
-wg use global sequence weighting for realignment!
Gap cost options:
-gapb [0,inf[ Transition pseudocount admixture (def=1.00)
-gapd [0,inf[ Transition pseudocount admixture for open gap (default=0.15)
-gape [0,1.5] Transition pseudocount admixture for extend gap (def=1.00)
-gapf ]0,inf] factor to increase/reduce gap open penalty for deletes (def=0.60)
-gapg ]0,inf] factor to increase/reduce gap open penalty for inserts (def=0.60)
-gaph ]0,inf] factor to increase/reduce gap extend penalty for deletes(def=0.60)
-gapi ]0,inf] factor to increase/reduce gap extend penalty for inserts(def=0.60)
-egq [0,inf[ penalty (bits) for end gaps aligned to query residues (def=0.00)
-egt [0,inf[ penalty (bits) for end gaps aligned to template residues (def=0.00)
Pseudocount (pc) options:
Context specific hhm pseudocounts:
-pc_hhm_contxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=2)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
3: CSBlast admixture: tau = a(1+b)/(Neff[i]+b)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_hhm_contxt_a [0,1] overall pseudocount admixture (def=0.9)
-pc_hhm_contxt_b [1,inf[ Neff threshold value for mode 2 (def=4.0)
-pc_hhm_contxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context independent hhm pseudocounts (used for templates; used for query if contxt file is not available):
-pc_hhm_nocontxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=2)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_hhm_nocontxt_a [0,1] overall pseudocount admixture (def=1.0)
-pc_hhm_nocontxt_b [1,inf[ Neff threshold value for mode 2 (def=1.5)
-pc_hhm_nocontxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context-specific pseudo-counts:
-nocontxt use substitution-matrix instead of context-specific pseudocounts
-contxt <file> context file for computing context-specific pseudocounts (default=${HHLIB}/data/context_data.crf)
-csw [0,inf] weight of central position in cs pseudocount mode (def=1.6)
-csb [0,1] weight decay parameter for positions in cs pc mode (def=0.9)
Other options:
-v <int> verbose mode: 0:no screen output 1:only warings 2: verbose (def=2)
-cpu <int> number of CPUs to use (for shared memory SMPs) (default=2)
-scores <file> write scores for all pairwise comparisons to file
-atab <file> write all alignments in tabular layout to file
-maxres <int> max number of HMM columns (def=20001)
-maxmem [1,inf[ limit memory for realignment (in GB) (def=3.0)
Example: hhsearch -i a.1.1.1.a3m -d scop70_1.71
Filter an alignment by maximum pairwise sequence identity, minimum coverage, minimum sequence identity, or score per column to the first (seed) sequence etc.
Usage: hhfilter -i infile -o outfile [options]
-i <file> read input file in A3M/A2M or FASTA format
-o <file> write to output file in A3M format
-a <file> append to output file in A3M format
Options:
-v <int> verbose mode: 0:no screen output 1:only warings 2: verbose
-id [0,100] maximum pairwise sequence identity (%) (def=90)
-diff [0,inf[ filter MSA by selecting most diverse set of sequences, keeping
at least this many seqs in each MSA block of length 50 (def=0)
-cov [0,100] minimum coverage with query (%) (def=0)
-qid [0,100] minimum sequence identity with query (%) (def=0)
-qsc [0,100] minimum score per column with query (def=-20.0)
-neff [1,inf] target diversity of alignment (default=off)
Input alignment format:
-M a2m use A2M/A3M (default): upper case = Match; lower case = Insert;
'-' = Delete; '.' = gaps aligned to inserts (may be omitted)
-M first use FASTA: columns with residue in 1st sequence are match states
-M [0,100] use FASTA: columns with fewer than X% gaps are match states
Example: hhfilter -id 50 -i d1mvfd_.a2m -o d1mvfd_.fil.a2m
Build a profile hidden Markov models from an input alignment, formatted in A2M, A3M, or FASTA, or convert between HMMER format (.hmm) and HHsearch format (.hhm).
Usage: hhmake -i file [options]
-i <file> query alignment (A2M, A3M, or FASTA), or query HMM
Output options:
-o <file> HMM file to be written to (default=<infile.hhm>)
-a <file> HMM file to be appended to
-v <int> verbose mode: 0:no screen output 1:only warings 2: verbose
-seq <int> max. number of query/template sequences displayed (def=10)
Beware of overflows! All these sequences are stored in memory.
-cons make consensus sequence master sequence of query MSA
-name <name> use this name for HMM (default: use name of first sequence)
Filter query multiple sequence alignment
-id [0,100] maximum pairwise sequence identity (%) (def=90)
-diff [0,inf[ filter MSA by selecting most diverse set of sequences, keeping
at least this many seqs in each MSA block of length 50 (def=100)
-cov [0,100] minimum coverage with query (%) (def=0)
-qid [0,100] minimum sequence identity with query (%) (def=0)
-qsc [0,100] minimum score per column with query (def=-20.0)
-neff [1,inf] target diversity of alignment (default=off)
Input alignment format:
-M a2m use A2M/A3M (default): upper case = Match; lower case = Insert;
'-' = Delete; '.' = gaps aligned to inserts (may be omitted)
-M first use FASTA: columns with residue in 1st sequence are match states
-M [0,100] use FASTA: columns with fewer than X% gaps are match states
Pseudocount (pc) options:
Context specific hhm pseudocounts:
-pc_hhm_contxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=0)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
3: CSBlast admixture: tau = a(1+b)/(Neff[i]+b)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_hhm_contxt_a [0,1] overall pseudocount admixture (def=0.9)
-pc_hhm_contxt_b [1,inf[ Neff threshold value for mode 2 (def=4.0)
-pc_hhm_contxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context independent hhm pseudocounts (used for templates; used for query if contxt file is not available):
-pc_hhm_nocontxt_mode {0,..,3} position dependence of pc admixture 'tau' (pc mode, default=2)
0: no pseudo counts: tau = 0
1: constant tau = a
2: diversity-dependent: tau = a/(1+((Neff[i]-1)/b)^c)
(Neff[i]: number of effective seqs in local MSA around column i)
-pc_hhm_nocontxt_a [0,1] overall pseudocount admixture (def=1.0)
-pc_hhm_nocontxt_b [1,inf[ Neff threshold value for mode 2 (def=1.5)
-pc_hhm_nocontxt_c [0,3] extinction exponent c for mode 2 (def=1.0)
Context-specific pseudo-counts:
-nocontxt use substitution-matrix instead of context-specific pseudocounts
-contxt <file> context file for computing context-specific pseudocounts (default=${HHLIB}/data/context_data.crf)
Example: hhmake -i test.a3m
Align a query alignment/HMM to a template alignment/HMM by HMM-HMM alignment. If only one alignment/HMM is given it is compared to itself and the best off-diagonal alignment plus all further non-overlapping alignments above significance threshold are shown. The command also allows to sample alignments randomly or to print out a list of indices of aligned residue pairs.
Usage: hhalign -i query -t template [options]
-i <file> input/query: single sequence or multiple sequence alignment (MSA)
in a3m, a2m, or FASTA format, or HMM in hhm format
-t <file> input/template: single sequence or multiple sequence alignment (MSA)
in a3m, a2m, or FASTA format, or HMM in hhm format
Input alignment format:
-M a2m use A2M/A3M (default): upper case = Match; lower case = Insert;
'-' = Delete; '.' = gaps aligned to inserts (may be omitted)
-M first use FASTA: columns with residue in 1st sequence are match states
-M [0,100] use FASTA: columns with fewer than X% gaps are match states
-tags/-notags do NOT / do neutralize His-, C-myc-, FLAG-tags, and trypsin
recognition sequence to background distribution (def=-notags)
Output options:
-o <file> write results in standard format to file (default=<infile.hhr>)
-oa3m <file> write query alignment in a3m or PSI-BLAST format (-opsi) to file (default=none)
-aa3m <file> append query alignment in a3m (-aa3m) or PSI-BLAST format (-apsi )to file (default=none)
-Ofas <file> write pairwise alignments in FASTA xor A2M (-Oa2m) xor A3M (-Oa3m) format
-add_cons generate consensus sequence as master sequence of query MSA (default=don't)
-hide_cons don't show consensus sequence in alignments (default=show)
-hide_pred don't show predicted 2ndary structure in alignments (default=show)
-hide_dssp don't show DSSP 2ndary structure in alignments (default=show)
-show_ssconf show confidences for predicted 2ndary structure in alignments
Filter options applied to query MSA, template MSA, and result MSA
-id [0,100] maximum pairwise sequence identity (def=90)
-diff [0,inf[ filter MSAs by selecting most diverse set of sequences, keeping
at least this many seqs in each MSA block of length 50
Zero and non-numerical values turn off the filtering. (def=100)
-cov [0,100] minimum coverage with master sequence (%) (def=0)
-qid [0,100] minimum sequence identity with master sequence (%) (def=0)
-qsc [0,100] minimum score per column with master sequence (default=-20.0)
-mark do not filter out sequences marked by ">@"in their name line
HMM-HMM alignment options:
-norealign do NOT realign displayed hits with MAC algorithm (def=realign)
-mact [0,1[ posterior prob threshold for MAC realignment controlling greedi-
ness at alignment ends: 0:global >0.1:local (default=0.35)
-glob/-loc use global/local alignment mode for searching/ranking (def=local)
Other options:
-v <int> verbose mode: 0:no screen output 1:only warings 2: verbose (def=2)
Example: hhalign -i T0187.a3m -t d1hz4a_.hhm -o result.hhr
Read one or many multiple alignments in one format and write them in another format
Usage: reformat.pl [informat] [outformat] infile outfile [options]
or reformat.pl [informat] [outformat] 'fileglob' .ext [options]
Available input formats:
fas: aligned fasta; lower and upper case equivalent, '.' and '-' equivalent
a2m: aligned fasta; inserts: lower case, matches: upper case, deletes: '-',
gaps aligned to inserts: '.'
a3m: like a2m, but gaps aligned to inserts MAY be omitted
sto: Stockholm format; sequences in several blocks with sequence name at
beginning of line (HMMER output)
psi: format as read by PSI-BLAST using the -B option (like sto with -M first -r)
clu: Clustal format; sequences in several blocks with sequence name at beginning
of line
Available output formats:
fas: aligned fasta; all gaps '-'
a2m: aligned fasta; inserts: lower case, matches: upper case, deletes: '-',
gaps aligned to inserts: '.'
a3m: like a2m, but gaps aligned to inserts are omitted
sto: Stockholm format; sequences in just one block, one line per sequence
psi: format as read by PSI-BLAST using the -B option
clu: Clustal format
If no input or output format is given the file extension is interpreted as format
specification ('aln' as 'clu')
Options:
-v int verbose mode (0:off, 1:on)
-num add number prefix to sequence names: 'name', '1:name' '2:name' etc
-noss remove secondary structure sequences (beginning with >ss_)
-sa do not remove solvent accessibility sequences (beginning with >sa_)
-M first make all columns with residue in first sequence match columns
(default for output format a2m or a3m)
-M int make all columns with less than X% gaps match columns
(for output format a2m or a3m)
-r remove all lower case residues (insert states)
(AFTER -M option has been processed)
-r int remove all lower case columns with more than X% gaps
-g '' suppress all gaps
-g '-' write all gaps as '-'
-uc write all residues in upper case (AFTER other options have been processed)
-lc write all residues in lower case (AFTER other options have been processed)
-l number of residues per line (for Clustal, FASTA, A2M, A3M formats)
(default=100)
-d maximum number of characers in nameline (default=1000)
Examples: reformat.pl 1hjra.a3m 1hjra.a2m
(same as reformat.pl a3m a2m 1hjra.a3m 1hjra.a2m)
reformat.pl test.a3m test.fas -num -r 90
reformat.pl fas sto '*.fasta' .stockholm
Add PSIPRED secondary structure prediction (and DSSP annotation) to a multiple sequence alignment (MSA) or HMMER (multi-)model file.
If the input file is an MSA, the predicted secondary structure and
confidence values are added as special annotation sequences with names
>ss_pred, >ss_conf, and >ss_dssp to the top of the output A3M
alignment. If no output file is given, the output file will have the
same name as the input file, except for the extension being replaced by
'.a3m'. Allowed input formats are A2M/FASTA (default), A3M (-a3m),
CLUSTAL (-clu), STOCKHOLM (-sto), HMMER (-hmm). Note that in order to
add PSIPRED and DSSP annotations corresponding paths have to be set in
HHPaths.pm. To add DSSP annotations addss.pl first checks a folder
of precomputed dssp files which can be obtained from
here. If the dssp file for a
particular structure is not available, addss.pl tries to execute the
DSSP binary mkdssp. Please keep in mind that only the current DSSP
version (2.2.1) supports structures in cif format, and that compiling
the software may cause problems on several systems. If you have troubles
to compile DSSP you may try to delete all occurrences of "static" in
its Makefile.
If the input file contains HMMER models, records SSPRD and SSCON containing predicted secondary structure and confidence values are added to each model. In this case the output file name is obligatory and must be different from the input file name.
Usage: perl addss.pl <ali file> [<outfile>] [-fas|-a3m|-clu|-sto]
or perl addss.pl <hmm file> <outfile> -hmm
From the top hits in an hhsearch output file (hhr), you can
-
generate a MSA (multiple sequence alignment) containing all representative template sequences from all selected alignments (options -fas, -a2m, -a3m, -pir)
-
generate several concatenated pairwise alignments in AL format (option -al)
-
generate several concatenated coarse 3D models in PDB format (option -ts)
In PIR, PDB and AL format, the pdb files are required in order to read the pdb residue numbers and ATOM records. The PIR formatted file can be used directly as input to the MODELLER homology modeling package.
Usage: hhmakemodel.pl [-i] file.hhr [options]
Options:
-i <file.hhr> results file from hhsearch with hit list and alignments
-fas <file.fas> write a FASTA-formatted multiple alignment to file.fas
-a2m <file.a2m> write an A2M-formatted multiple alignment to file.a2m
-a3m <file.a3m> write an A3M-formatted multiple alignment to file.a3m
-m <int> [<int> ...] pick hits with specified indices (default='-m 1')
-p <probability> minimum probability threshold
-e <E-value> maximum E-value threshold
-q <query_ali> use the full-length query sequence in the alignment
(not only the aligned part);
the query alignment file must be in HHM, FASTA, A2M,
or A3M format.
-N use query name from hhr filename (default: use same
name as in hhr file)
-first include only first Q or T sequence of each hit in MSA
-v verbose mode
Options when database matches in hhr file are PDB or SCOP sequences
-pir <file.pir> write a PIR-formatted multiple alignment to file.pir
-ts <file.pdb> write the PDB-formatted models based on *pairwise*
alignments into file.pdb
-al <file.al> write the AL-formatted *pairwise* alignments into file.al
-d <pdbdirs> directories containing the pdb files (for PDB, SCOP, or DALI
sequences)
-s <int> shift the residue indices up/down by an integer
-CASP formatting for CASP (for -ts, -al options)
(default: LIVEBENCH formatting)
Analogously, hhmakemodel.py generates a MSA containing all
representative template sequences from all selected alignments in PIR
format. Note that in hhmakemodel.py you have to specify the folder
containing the *.cif files which make up the alignment. The script will
modify the ATOM section of those cif files according to the residue
numbering in the alignment. Use these renumbered cifs file together with
the PIR alignment output in
MODELLER.
usage: hhmakemodel.py [-h] [-v] [-m INT [INT ...]] [-e FLOAT] [-r FLOAT] [-c]
FILE DIR FILE DIR
Creates a MODELLER alignment (*.pir) from a HHSearch results file (*.hhr).
positional arguments:
FILE results file from HHsearch with hit list and alignment
DIR path to the folder containing cif files
FILE output file (PIR-formatted multiple alignment)
DIR path to the folder where modified cif files should be
written to
optional arguments:
-h, --help show this help message and exit
-v, --verbose verbose mode
-m INT [INT ...] pick hits with specified indices (e.g. -m 2 5)
-e FLOAT maximum E-Value threshold (e.g. -e 0.001)
-r FLOAT residue ratio (filter alignments that have contribute at
least residues according to the specified ratio).
-c convert non-canonical residues (default = True)
Builds HHsuite database from a3m formatted MSAs and/or from HMMs (-o). MSAs and HMMs can also be added (-a) to or removed (-r) from an existing database.
Usage: hhsuitedb.py [options]
Options:
-h, --help show this help message and exit
--ia3m=<GLOB> Glob for a3m files
--ics219=<GLOB> Glob for cs219 files
--ihhm=<GLOB> Glob for hhm files
-o FILE Output hhsuite database basename
--cpu=INT Number of threads that shall used
--force Try to fix problems with the database
Write files into current directory and name each file by the first word after ">" in the name line.
Usage: splitfasta.pl infile [option]
Option:
-fam : use family-based name (for SCOP/ASTRAL sequences
-name : use sequence name as file name (default)
-ext <ext> : extension for sequence files (default=seq)
The program reads an input sequence in FASTA/A3M which must have a SCOP domain, PDB chain, or DALI
domain identifier (e.g. d1hz4a_, 1hz4_A, or 1hz4A_1). It reads the corresponding PDB file from the
given pdb directory and generates a new PDB file by aligning the input sequence to the sequence
extracted from the ATOM records of the corresponding pdb chain and renumbering the residues in
columns 23-26 according to the position in the input sequence.
(HETATM records for MSE (selenomethionine) will be interpreted as ATOM records for MET in the
alignment. MSE will be changed to MET in the output file.)
Usage: renumberpdb.pl [options] infile [outfile]
Example: renumberpdb.pl d1hz4a_.a3m /cluster/tmp/d1hz4a_.pdb
Options:
-o <file> output file (default: <infile_wo_extension>.pdb
-d <pdbdir> give directory of pdb files (default=)
-a [A|B] filter alternative location identifier (e.g. A or B)
An example of the usage of cif2fasta.py is provided in Building customized databases.
Usage: cif2fasta.py -i cif_folder -o *.fasta -c num_cores -v
cif2fasta.py takes a folder that contains cif files as input and outputs their
sequences into fasta file.
Options:
-h, --help show this help message and exit
-i DIR input cif folder.
-o FILE output fasta file.
-p FILE output PDB filter file (optional).
-s FILE SCOP annotation.
-c INT number of cores (default = 1).
-l INT Remove chains with a length < X (default = 30).
-v INT Verbose Mode (quiet = 0, full verbosity = 2).
An example of the usage of pdbfilter.py is provided in Building customized databases. Note that the annotations file which is required
to select the proper PDB is created by cif2fasta.py using the -p
flag.
usage: pdbfilter.py [-h] [-i FILE] [-r FILE] [-v] FILE FILE FILE FILE
pdbfilter.py selects from sequence clusters (determined by MMseqs2) the
sequences which have the best resolution, R-free factor and/or completness and
writes them to a fasta file.
positional arguments:
FILE input fasta file (created by cif2fasta.py)
FILE sequence clusters (MMseqs)
FILE annotations file (created by cif2fasta using the -p
flag, contains information about resolution, R-free
and completness of sequences).
FILE output fasta file
optional arguments:
-h, --help show this help message and exit
-i FILE, --include FILE
include PDB chains
-r FILE, --remove FILE
exclude PDB chains
-v, --verbose verbose mode
The HHsuite software package is distributed under Gnu Public License, Version 3.
This means that the HHsuite is free software: you can redistribute it and/or modify it under the terms of the GNU General Public License as published by the Free Software Foundation, either version 3 of the License, or (at your option) any later version.
This program is distributed in the hope that it will be useful, but WITHOUT ANY WARRANTY; without even the implied warranty of MERCHANTABILITY or FITNESS FOR A PARTICULAR PURPOSE. See the GNU General Public License for more details.
You should have received a copy of the GNU General Public License along
with this program in the file LICENSE. If not, see
http://www.gnu.org/licenses/.
- Johannes Söding: main code base incl. perl scripts up to version 2.0, debugging, documentation. Contact at soeding@mpibpc.mpg.de
- Milot Mirdita: Uniclust30 database and maintenance
- Markus Meier: uniprot-1it database and associated hhblits code, code refactoring, tests and benchmarks
- Martin Steinegger: Fast SIMD HMM-HMM Viterbi code and various speed-ups
- Michael Remmert: early HHblits version
- Andy Hauser: ffindex, speed/memory optimization
- Christof Angermüller: context-specific pseudocounts, speed optimizations
- Armin Meier: HHblits column state alphabets, tests and benchmarks
- Andreas Biegert: HHblits column state alphabet cs219, context-specific pseudocounts
We thank the many users of HHsuite who sent bug reports or gave critical feedback that led to improvements in the software. We are grateful to Lazlo Kajan, Stefan Möller and the Debian team for making the HHsuite software available as a Debian package. J.S. thanks Andrei Lupas for his invaluable support, encouragement, and feedback during the first five years of the work on HHsuite in his department.
For many years, the HH-suite contained code adapted from Michael Farrar for its Smith-Waterman implementation. We want to express our posthumous gratitude to Michael Farrar for his code. The current Smith-Waterman implementation is based on the SSW library from Mengyao Zhao, for which we are very grateful.
Reference:
-
Farrar M (2007). Striped Smith-Waterman speeds database searches six times over other SIMD implementations. Bioinformatics 23, 156-161. doi: 10.1093/bioinformatics/btl582
-
Zhao M, Lee W P, Garrison E P, Marth G T (2013). SSW Library: An SIMD Smith-Waterman C/C++ Library for Use in Genomic Applications. PLOS One 8, e82138. doi: 10.1371/journal.pone.0082138
Good luck with your work!